

- Crossoptilon : le 4 specie ( & sottospecie - ibridi ecc., )... tra mito e realtà -
Da sinistra, sotto Crossoptilon mantchuricum - Crossoptilon harmani coppia con prole sopra - a destra Crossoptilon auritum sotto - Crossoptilon crossoptilon sopra

Crossoptilon specie e sottospecie
Crossoptilon – krossós = frangia – fringe; villus = vello – pelt; ptílon = piuma – feather; ala = ala – wing
"..Le attuali informazioni, nonché il buon senso che non dovrebbe avere epoca, suggeriscono che in questo genere potrebbero essere incluse, perlomeno, due forme non meritevoli dello status specifico in quanto sia per il Crossoptilon c. dolani che lichiangense, l’origine ibrida sembra plausibile. Se per il primo persiste il dubbio che, perlomeno agli albori, potesse rappresentare una mutazione leggermente più scura della forma nominale, nel caso del Crossoptilon lichiangense, a fronte sia della colorazione, che della distribuzione letteralmente incastonata tra quelle dei congenere crossoptilon e drouyni, la condizione ibrida parrebbe verosimile. Nel caso in cui "originariamente" il Crossoptilon c. dolani fosse stata una variante del Crossoptilon c. crossoptilon, attualmente sarebbe alquanto..."

"..difficile separalo dalla moltitudine di fenotipi, sicuramente ibridi, in tutte le gradazioni di grigio, che vivono sia ai limiti, quanto nella gamma storica attribuita a questa forma. Quindi è auspicabile che il suo status venga rivisto e, se necessario riconsiderato, nonché adattato ad una eventuale nuova condizione.
A tale proposito, l'esaminazione di una notevole quantità di immagini, con località indicate, parrebbe suggerire che l'unica specie compresa in questo genere ad essere realmente isolata è il Crossoptilon mantchuricum. Mentre le gamme delle altre si incontrano e sovrappongono. Nel caso dei Crossoptilon c. drouyni e dolani, addirittura, parrebbe non esserci più alcuna separazione in totale promiscuità territoriale.."
Ai due sopra menzionati, aggiungerei anche il Crossoptilon c. drouyni la cui posizione intermedia, tra la forma nominale ed il non riconosciuto Crossoptilon c. leucurum, suggerisce alcuni dubbi sulla sua posizione specifica. Tuttavia, non si può pretendere di giudicare il presente senza conoscere, quand’anche solo per sommi capi, il passato con particolare riferimento a ciò che argomentavano ornitologi e studiosi in riguardo a queste forme. Quindi, a tale proposito, oltre a studi recenti che chiariscono, o perlomeno provano a farlo, alcuni aspetti rimasti a lungo irrisolti di questi stupendi quanto intriganti galliformi, ritengo interessante proporre i lavori pubblicati da autori del passato, incluse, se possibile, le descrizioni originali delle specie comprese in questo genere.
Una sorta di breve cronaca temporale che mette in evidenza sia la confusione iniziale riguardante al classificazione di alcune forme che l'interesse per questo genere da parte degli ornitologi. Non una disamina vera e propria, dunque, ma una attenta panoramica delle specie da semplice appassionato.
Alessandro Ghigi nel suo interessantissimo studio: "Ricerca Ibridologica si Crossoptilon" consultabile a fondo pagine e che consiglio di leggere, in particolare la seconda parte, considera il Crossoptilon c. leucurum specificatamente distinto e drouyni un intermedio. A seguito un brevissimo sunto del lavoro citato:
"...Gli esemplari del Museo di Parigi confermano i dati di Lord Rothschild: tutti i leucurum sono di colore bianco puro, ad eccezione delle punte delle timoniere, dei rachide delle primarie e della sommità della testa, che sono neri. Crossoptilon (tibetanum) presenta una peluria basale sulle timoniere posteriori, lati sempre più scure nelle parti coperte delle timoniere fino alle remiganti e timoniere caudali interamente blu con una sfumatura acciaio. C. harmani assomiglia all'auritum per colore, ma ha le parti inferiori bianche e una coda acciaio, macchiata di bianco lungo la parte interna della membrana esterna delle timoniere laterali.
Se volessimo rappresentare la gradazione delle tonalità in ordine lineare, dal bianco al blu e al marrone, dovremmo disporre la serie come segue:
leucurum (Seeb.) drouyni (Verr.) crossoptilon (Hodgs) harmani (Elwes.) auritum (Pall.) -- mantchuricum (Swinb.)
Le prime quattro forme sono ovviamente le più primitive: probabilmente la specie originaria è crossoptilon, che si è evoluta a ovest in leucurum e a est in harmani, mentre auritum e mantchuricum sono due forme più distinte e notevolmente divergenti. Infatti, nei giovani di queste ultime forme, quando sono nel loro primo piumaggio, sono presenti solo 20 penne della coda con barbe normali; la muta è centripeta come in tutti i fagiani; sono le penne laterali della coda a cambiare per prime. Quelle caudali mediane con barbe decomposte compaiono contemporaneamente alle laterali; rappresentano quindi un'acquisizione tardiva che non ha corrispondenza in altri fagiani. I ciuffi auricolari nei giovani non si estendono oltre la testa. Pertanto, in auritum e mantchuricum, sono presenti due fattori di sviluppo: penne mediane della coda decomposte e ciuffi auricolari lunghi, che non appartengono al gruppo crossoptilon-leucurum-harmanti.
Lo stato intermedio degli ibridi dimostra inoltre che auritum e mantchurieum si sono evoluti in uno stato di isolamento geografico, mentre crossoptilon è sempre stato in comunione geografica con i suoi stretti alleati, come dimostrato dai numerosi intermedi rinvenuti in natura, di cui drouyni è il più noto.
Speriamo di importare esemplari vivi di Crossoptilon crossoptilon (tibetanum); l'incrocio con auritum potrebbe risolvere i dubbi rimanenti sulle affinità naturali delle diverse forme di questi curiosi fagiani..."
- "..Il Crossoptilon, appartenente alla famiglia dei Phasianidae nell'ordine dei Galliformi, è un genere raro ma importante ed endemico in Cina comprendente quattro specie tra cui il fagiano orecchiuto bianco (Crossoptilon crossoptilon), il fagiano orecchiuto blu (Crossoptilon auritum), il fagiano orecchiuto bruno (Crossoptilon mantchuricum) ed il fagiano orecchiuto tibetano (Crossoptilon harmani). Prediligono albergare nelle foreste di conifere e miste con latifoglie e conifere nonché le aree arbustive alpine in varie parti della Cina ad altitudini molto diverse che vanno da 20 m., a 5.000 m. Si ritiene che il Crossoptilon crossoptilon e harmani siano conspecifici e abitino simpatricamente la foresta pluviale ad altitudini elevate di 3.000-5.000 m. Crossoptilon harmani è distribuito nel Tibet sudorientale e nell'adiacente India settentrionale (3.000-5.000 m) 1*), mentre Crossoptilon crossoptilon vive nel Qinghai, Sichuan, Yunnan e Tibet (3.000-4.300 m). Crossoptilon auritum e mantchuricum risultano strettamente correlati, ma abitano allopatricamente aree diverse. Il primo si incontra solo sulle montagne del Qinghai, Gansu, Sichuan e Ningxia, a un'altitudine intermedia di 1.500-3.000 m., mentre il bruno è distribuito principalmente nelle montagne dello Shanxi, dell'Hebei e nei pressi di Pechino, a un'altitudine bassa di 20-1.000 m.."
- "..Elliot li considerava prossimi a Lophophorus e Pavo, a causa delle mandibole, forti arti e disintegrazione del piumaggio. Altri autori lo collocavano trai Lobiophasis, probabilmente per il numero insolitamente elevato di penne della coda.."
- "..Mostrano una colorazione specifica che, in generale, può essere riassunta in bianca, blu e marrone, con penne della coda parzialmente iridescenti e semi-disintegrate in numero di venti-ventiquattro nell'adulto, tutte grandi, larghe e arrotondate. Quelle centrali pendenti, con estese membrane libere, lunghe il doppio della coppia esterna.
La prima primaria è molto più corta della seconda, che è lunga quanto l'ottava o la decima; la quinta o la sesta è la più lunga della serie.
La sommità del capo è ricoperta di piume corte, morbide e arricciate, dalla consistenza vellutata. Le copritrici auricolari sono molto allungate, perlomeno nelle forme blu e marrone, modellando lunghi ciuffi bianchi di piume piuttosto rigide, a volte ricurvi verso l'alto, su ciascun lato del capo. Il resto del piumaggio è denso, lungo e piuttosto "peloso".
I lati della faccia sono nudi, rossastri e ricoperti di piccole papille. I sessi sono simili, tranne per il fatto che i maschi oltre ad essere solitamente più grandi, sono dotati di speroni corti e robusti. Il tarso è considerevolmente più lungo del dito medio e dell'unghia.
Le tonalità brunastre degli uccelli immaturi si ritrovano nel piumaggio adulto di una specie, mentre in un'altra questa tonalità si trasforma in un blu ardesia. Questo a sua volta cede sempre più il passo al bianco, fino a raggiungere l'apice dell'evoluzione cromatica nel fagiano orecchiuto tibetano quasi completamente bianco.."
Commento personale: 1*) - Birdlife nel 2024 esclude che qualche forma di Crossoptilon si possa trovare nelle aree indiane e birmane (Myanmar). Vedere alla parte dedicata al Crossoptilon crossoptilon.
Crossoptilon ( Hodgson B.H. ... 1838 )
Crossoptilon auritum..................fagiano orecchiuto blu.................................Pallas P.S. .............................1811
Crossoptilon c. crossoptilon......fagiano orecchiuto bianco...........................Hodgson B.H. ......................1838
* Crossoptilon c. drouynii..............fagiano orecchiuto bianco di Drouyn..........Verreaux J.P. .......................1868
* Crossoptilon c. dolani.................fagiano orecchiuto "bianco" di Dolan.........Meyer de Schauensee R. ....1937
* Crossoptilon c. lichiangense......fagiano orecchiuto bianco di Lijiang...........Delacour J.T. ........................1945
Crossoptilon mantchuricum......fagiano orecchiuto bruno.............................Swinhoe R. ...........................1863
Crossoptilon harmani.................fagiano orecchiuto di Harman......................Elwes H.J. ............................1881
I risultati di uno studio filogenetico condotto nel 2014 indicano che il genere Crossoptilon era fratello del genere Lophura.
Mappa della distribuzione allegata alle note di Delacour del 1945 esposta per esteso in questa "panoramica".

Questo genere occupa un vasto areale che indicativamente si propaga dalla catena montuosa degli Alashan, compresi tra le province cinesi del Ningxia e Inner Mongolia, che parrebbero segnarne il limite settentrionale, a raggiungere le aree del Tibet sud-orientale sino ai confini dell'Arunachal Pradesh (India) ed ancora più a sud nella provincia dello Yunnan (Cina) perlomeno sino alla prefettura di Lijiang o Likiang.
Elliot li considerava prossimi a Lophophorus e Pavo, a causa delle mandibole, forti arti e disintegrazione del piumaggio. Altri autori lo collocavano trai Lobiophasis, forse per il numero insolitamente elevato di penne della coda.
Mostrano una colorazione specifica che, in generale, può essere riassunta in bianca, blu e marrone, con penne della coda parzialmente iridescenti e semi-disintegrate in numero di venti-ventiquattro nell'adulto, tutte grandi, larghe e arrotondate. Quelle centrali pendenti, con estese membrane libere, lunghe il doppio della coppia esterna.
La prima primaria è molto più corta della seconda, che è lunga quanto l'ottava o la decima; la quinta o la sesta è la più lunga della serie.
La sommità del capo è ricoperta di piume corte, morbide e arricciate, dalla consistenza vellutata.
Le copritrici auricolari sono molto allungate, perlomeno nelle forme blu e marrone, modellando lunghi ciuffi bianchi di piume piuttosto rigide, a volte ricurvi verso l'alto, su ciascun lato del capo. Il resto del piumaggio è denso, lungo e piuttosto "peloso".
I lati della faccia sono nudi, rossastri e ricoperti di piccole papille. I sessi sono simili, tranne per il fatto che i maschi oltre ad essere solitamente più grandi, sono dotati di speroni corti e robusti. Il tarso è considerevolmente più lungo del dito medio e dell'unghia.
Le tonalità brunastre degli uccelli immaturi si ritrovano nel piumaggio adulto di una specie, mentre in un'altra questa tonalità si trasforma in un blu ardesia. Questo a sua volta cede sempre più il passo al bianco, fino a raggiungere l'apice dell'evoluzione cromatica nel fagiano orecchiuto tibetano quasi completamente bianco.
Foto da: 1 Autore: Lin Hillside - 2 www.birdnet.cn - 3 www.ebird.org Autori: Lisa & Li Li - 4 www.birdnet.cn Autore: Village chief
1 - Crossoptilon auritum 2 - Crossoptilon c. crossoptilon
.jpg)
_edit.jpg)
3 - Crossoptilon mantchuricum 4 - Crossoptilon harmani
%2C%20Shanxi%2C%20China%2019-05-25%20-%20Lisa%20%26%20Li%20Li%20ebird%20org_PNG.png)

Studio pubblicato nel 2000, quindi antecedente al precedente, che mette soprattutto i risalto la variazione all'interno di alcune forme:
Bird Conservation International (2000) © BirdLife International 2000
Perché i fagiani orecchiuti dell'altopiano orientale del Qinghai-Tibet presentano così tante variazioni morfologiche? XIN LU e GUANG-MEI ZHEN
Riepilogo È noto che i fagiani orecchiuti bianchi (Crossoptilon crossoptilon drouyni) si incrociano ampiamente con i fagiani orecchiuti tibetani (Crossoptilon harmani) ai confini dei loro areali. Una nuova zona ibrida è stata recentemente scoperta nel Tibet orientale, ben lontana dai confini degli areali delle specie parentali. Sulla base di osservazioni ecologiche sui fagiani dalle orecchie bianche e della storia geografica e della struttura dei ghiacciai moderni, abbiamo attribuito la complessa variazione morfologica dei fagiani dalle orecchie bianche e l'elevata biodiversità dell'altopiano orientale del Qinghai-Tibet alla sua geografia variegata.
Introduzione Il genere di fagiano orecchiuto (Crossoptilon) è endemico della Cina e comprende quattro specie. Il fagiano orecchiuto marrone C. mantchuricum si trova nella Cina settentrionale e il fagiano orecchiuto blu C. auritum si trova sull'altopiano del Qinghai Tibet settentrionale. Queste specie non mostrano variazioni morfologiche e non hanno sottospecie descritte. Il fagiano orecchiuto bianco C. crossoptilon mostra maggiori variazioni nella colorazione del piumaggio e sono state riconosciute quattro sottospecie: Crossoptilon nel Sichuan occidentale e nel Tibet sudorientale, lichiangense nello Yunnan nordoccidentale, drouyni nell'area tra il fiume Nujiang e il fiume Jinsha e dolani nello Yushu del Qinghai meridionale. Il fagiano orecchiuto tibetano C. harmani è limitato al Tibet, a nord dell'asse principale dell'Himalaya (Ludlow e Kinnear 1944). In passato era considerato una sottospecie del fagiano orecchiuto bianco (Delacour 1977, Cheng et al. 1978), ma più di recente è stato considerato una specie a tutti gli effetti (Sibley e Monroe 1990, Cheng 1994) 1*), principalmente per il suo piumaggio grigio-blu scuro, che si distingue da quello prevalentemente bianco delle altre sottospecie. Le popolazioni tibetane presentano una colorazione del piumaggio molto uniforme in gran parte dell'areale. Tuttavia, al confine nord-orientale del loro areale, gli uccelli si incrociano con i drouyni e sono stati registrati alcuni ibridi (Delacour 1945, Cheng et al. 1978) 2*). Variazioni più enigmatiche nel piumaggio di questi fagiani sono state riscontrate in altre aree dell'altopiano orientale del Qinghai-Tibet. All'interno dell'areale dei Crossoptilon drouyni, sono stati trovati individui con macchie bianche sulla sommità nera della testa, mentre sono stati registrati uccelli con penne della coda parzialmente o interamente bianche (Cheng et al. 1983) 3*). Esistono modelli di piumaggio ancora più complessi nel Tibet nord-orientale e Qinghai meridionale dove si incontrano gli intervalli di Crossoptilon crossoptilon, drouyni e dolani (Pack-Blumenau e Lu 1999, X. Lu oss. pers.). Considerato che questa variazione nei fagiani orecchiuti è stata registrata anche in altri uccelli della regione si ritiene che possa essere attribuita alla complessità geografica del vasto altopiano orientale (Tang et al. 1996, Lu et al. 1998). La comprensione di questi modelli di variazione è importante non solo per gli studi evolutivi e sistematici, ma anche per la conservazione (Pojas 1992). Tuttavia, non è stato ancora fornito un resoconto dettagliato di come le condizioni geografiche responsabili della limitazione dei flussi genici tra le popolazioni abbiano portato alla maggiore diversità di specie nell'altopiano orientale.
Osservazioni Nel maggio 1995, abbiamo scoperto un gruppo di ibridi tra C. c. drouyni e C. harmani in una località chiamata Sawang, nel Tibet orientale (93°39'E, 32°24'N), non precedentemente visitata dai biologi. Le seguenti caratteristiche di questi ibridi si basano su osservazioni sul campo e su due esemplari adulti, oltre a diverse mute di piume (tra cui 91 piume della coda, 41 delle ali e 488 altre piume). Gli uccelli mostravano un piumaggio bianco-blu, una combinazione tra il bianco puro tipico dei drouyni e il blu scuro tipico degli harmani. Molte delle loro piume primarie erano scure, derivanti dagli harmani, con alcune piume completamente bianche o con macchie bianche, come i drouyni. In questi ultimi la maggior parte della coda è bianca, sfumando in un blu-verde scuro verso la punta; negli harmani, è grigiastra vicino alla base della coda, mentre il resto è di un blu nero metallico. Ma le basi delle penne della coda degli ibridi erano spesso macchiate di bianco. Un giovane esemplare (di circa tre mesi) pesava 1,18 kg e mostrava un piumaggio simile a quello dell'ibrido adulto. Le uova ibride differivano da quelle dei genitori per colore e dimensioni. La lunghezza e larghezza medie di nove uova ibride erano rispettivamente di 55,1 mm (DS ± 1,73) e 41,2 mm (± 1,02), significativamente inferiori a quelle di sei uova di drouyni (lunghezza 59,1 mm ± 1,46, t = 4,56, P < 0,001; larghezza 43,5 mm ± 1,43, t = 3,39, P < 0,005); ma più simili a quelle di 40 uova di harmani (lunghezza 57,3 mm ± 3,38, t = 1,72, P = 0,09, larghezza 41,7 mm ± 1,01, t = 1,29, P = 0,20). I pulcini ibridi appena nati presentavano un piumaggio simile a quello dei genitori. Topograficamente, Sawang comprende alte montagne e profonde valli, con una vegetazione naturale stratificata verticalmente. Due tipi di foresta sui pendii esposti a sud costituiscono gli habitat preferiti dagli ibridi: la quercia a foglie di agrifoglio Quercus aquifolioides a quote più basse (3.700-4.300 m) e il ginepro tibetano Sabina tibetica più in alto (4.200-4.700 m) (Lu 1997). Il 3 agosto 1995, lasciammo Sawang ed esplorammo la valle del fiume Niwuzangbu, incontrando spesso ibridi. Il giorno dopo, risalimmo il ghiacciaio Aigagong a 5.400 m e entrammo nella valle successiva. Scendendo nella foresta di ginepri tibetani, con nostra sorpresa scoprimmo che i fagiani avevano un piumaggio grigio-bluastro ed erano harmani puri. I fagiani orecchiuti sui lati opposti della cresta montuosa non potevano incontrarsi, perché le nostre osservazioni sull'uso dell'habitat da parte degli ibridi confermavano che non si erano mai spinti oltre i 100 m sopra il limite della vegetazione arborea (4.800 m di altitudine). Come si producono, quindi, questi ibridi nella nostra area di studio? In altre parole, come si incrociano le specie parentali, C. c. drouyni e C. harmani ? Se esaminiamo la topografia e la struttura glaciale su un areale più ampio (Figura 1) Si può osservare che il fiume Yigongzangbu (in cui confluisce il fiume Niwuzangbu) attraversa alte montagne per entrare nella valle del fiume Yaluzangbu. Qui, si raccoglie aria calda e umida proveniente dall'Oceano Indiano e si formano ghiacciai marittimi nella parte centrale dei monti Nianqingtanghgula. La valle dello Yigongzangbu si apre verso un'ansa del fiume Yaluzangbu, formando così un canale unico attraverso il quale l'aria calda e umida penetra nel Tibet settentrionale (Yang et al. 1983). A giudicare da precedenti raccolte (ad esempio Delacour 1945, Cheng et al. 1983) e dai dati di questa indagine, drouyni e harmani sono presenti su entrambe le sponde del medio corso del fiume Yigongzangbu e vi è anche una registrazione di entrambe le specie in simpatria a Pomi conosciuta soprattutto come Bōmì o Bomê (Figura 1). È probabile che l'ibridazione tra queste specie sia avvenuta nel punto in cui i loro areali si incontravano. In competizione con i loro genitori, gli ibridi furono costretti a cercare nuovi territori. Essendo scarsi volatori, non potevano attraversare le alte montagne e i ghiacciai nelle aree circostanti e dovettero disperdersi lungo la valle dello Yigong-zangbu negli affluenti del fiume, inclusa la valle del Niwuzangbu (Figura 1). In effetti, abbiamo osservato e siamo stati anche informati (dal Professor Gu del Tibet Plateau Institute of Biology) che gli ibridi si trovano lungo la valle principale dello Yigong-zangbu
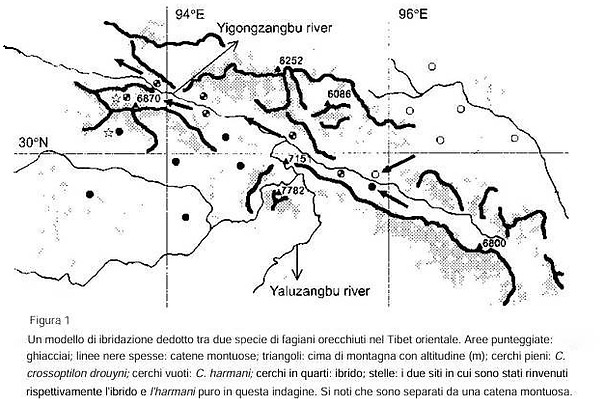
Discussione Una zona ibrida è una regione di forte integrazione genetica e fenotipica tra popolazioni contigue geneticamente distinte e relativamente uniformi (Mayr 1963). Sono state documentate oltre 170 zone ibride ragionevolmente chiare tra piante e animali (Barton e Hewitt 1989), la maggior parte delle quali si verificano dove si trovano i loro genitori.
Le specie si incontrano direttamente. La complessa dispersione dell'ibridazione nei fagiani orecchiuti è particolarmente interessante. Il fatto che questa popolazione ibrida possa riprodursi con successo e sia una componente predominante della comunità galliforme locale indica che gli ibridi sono vitali e potrebbero essere stati stabiliti nella zona per lungo tempo. Sotto il concetto di specie biologica (Mayr 1963), harmani non può essere una singola specie. Tuttavia, sulla base del concetto di specie filogenetica (Cracraft 1983), i due geni parentali i taxa dovrebbero essere considerati come specie distinte che hanno mantenuto la capacità di incrociarsi (Zink e McKitrick 1995). Ricerca completa sulla storia naturale dell'altopiano del Qinghai-Tibet rivela che l'altopiano ha avuto un processo evolutivo molto breve (Li et al. 1986).
Durante tre milioni di anni dal tardo Terziario al tardo Quaternario, il le montagne dell'altopiano orientale si elevavano fino a 3.000 m di altitudine (da 1.000 a oltre 4.000 m), gran parte di questo cambiamento si è verificato forse solo 100.000 anni fa. Questi cambiamenti ambientali del Pleistocene hanno avuto un impatto importante sulla diversificazione genetica microevolutiva aviaria (Avise e Walker 1998). La rapida e drastica cambiamenti geologici nell'altopiano tibetano orientale, insieme ad un'alternanza tra le epoche glaciali e interglaciali a partire dal Quaternario deve essere stato notevolmente ha interessato la speciazione nel genere Crossoptilon, che probabilmente ha avuto origine dal Himalaya orientale nel Terziario (Lu et al. 1998). Piccola scala geografica I cambiamenti potrebbero dare inizio alla speciazione tra questi uccelli che volano male e non migrano fagiani. È la contraddittoria coesistenza di barriere geografiche e corridoi fluviali che ha prodotto la diversità aviaria nell'area che osserviamo oggi. Finora, l'ipotesi non è stata verificata. Sono previste ulteriori indagini per raccogliere ulteriori dati sulla variazione morfologica e molecolare dei fagiani orecchiuti, così come ulteriori dati sull'ecologia e sul comportamento, in tutto il loro areale. Ciò consentirà ci permettono di valutare l'ibridazione e il flusso genico in questi uccelli. Sulla base dei cambiamenti storici e degli attuali modelli ambientali dell'altopiano, potremmo essere in grado di spiegare la vera causa di questa enigmatica variazione e chiarire l'evoluzione dei fagiani dalle orecchie lunghe in quest'area chiave per la conservazione della natura in Asia
A seguire un interessantissimo studio condotto nel 2015:
BMC Genomics. 2015 febbraio 5;16(1):42. doi: 10.1186/s12864-015-1234-9
Genomica mitocondriale comparativa e relazioni filogenetiche della specie Crossoptilon (Phasianidae, Galliformes)
Xuejuan Li, Yuan Huang, Fumin Lei .
Codice PMC: PMC4326528 Codice PMI: 25652939
Testo per esteso, figure e file aggiuntivi consultabili al seguente link: https://pmc.ncbi.nlm.nih.gov/articles/PMC4326528/#Sec20
- Materiale supplementare elettronico
La versione online di questo articolo (doi:10.1186/s12864-015-1234-9) contiene materiale supplementare, disponibile agli utenti autorizzati.
Parole chiave: Genoma mitocondriale, Crossoptilon , Filogenesi, Tempo di divergenza, Ka/Ks
- Astratto
"..I Phasianidae costituiscono una famiglia di Galliformi composta da 38 generi e circa 138 specie, raggruppati in due tribù in base alle loro caratteristiche morfologiche, i Fagiani e le Pernici. Diversi studi hanno tentato di ricostruire le relazioni filogenetiche dei Phasianidae, ma molti interrogativi rimangono ancora irrisolti, come lo status tassonomico e le relazioni filogenetiche tra le specie di Crossoptilon. Il genoma mitocondriale (mitogenoma) è stato ampiamente utilizzato per dedurre la diversificazione genetica aviaria con una risoluzione ragionevole. In questo studio, abbiamo sequenziato l'intero mitogenoma di tre specie di Crossoptilon (C. harmani , C. mantchuricum e C. crossoptilon) per indagare la loro relazione evolutiva.."
- Risultati
"..I mitogenomi completi di C. harmani , C. mantchuricum e C. crossoptilon sono rispettivamente lunghi 16682 bp, 16690 bp e 16680 bp e codificano un set standard di 13 geni codificanti proteine, 2 geni dell'RNA ribosomiale, 22 geni dell'RNA di trasferimento e una presunta regione di controllo. C. auritum e C. mantchuricum sono geneticamente più correlati, mentre C. harmani è più strettamente associato a C. crossoptilon. Crossoptilon mostra una relazione più stretta con Lophura ed è stata ricostruita la seguente relazione filogenetica: (( Crossoptilon + Lophura ) + ( Phasianus + Chrysolophus )). Il tempo di divergenza tra i cladi C. harmani - C. crossoptilon e C. mantchuricum - C. auritum è coerente con il sollevamento dell'altopiano tibetano durante il Pliocene terziario. L'analisi Ka/Ks ha mostrato che il gene atp8 nel Crossoptilon ha probabilmente subito una forte pressione selettiva nell'adattamento all'ambiente dell'altopiano.."
- Conclusioni
"..C. auritum con C. mantchuricum e C. harmani con C. crossoptilon formano due coppie di gruppi fratelli. La distanza genetica tra C. harmani e C. crossoptilon è di gran lunga inferiore alla distanza interspecifica ed è prossima alla distanza intraspecifica di Crossoptilon, indicando che C. harmani è molto più strettamente imparentato con C. crossoptilon. La nostra analisi mito-filogenomica supporta la monofilia di Crossoptilon e la sua più stretta relazione con Lophura. Si suggerisce che il sollevamento dell'altopiano tibetano abbia influenzato la divergenza tra il clade C. harmani - C. crossoptilon e il clade C. mantchuricum - C. auritum durante il Pliocene terziario. Il gene Atp8 nella specie Crossoptilon potrebbe aver subito una forte pressione selettiva per l'adattamento all'ambiente dell'altopiano.."
- Materiale supplementare elettronico
"..La versione online di questo articolo (doi:10.1186/s12864-015-1234-9) contiene materiale supplementare, disponibile agli utenti autorizzati.
Parole chiave: Genoma mitocondriale, Crossoptilon , Filogenesi, Tempo di divergenza, Ka/Ks.."
Crossoptilon, appartenente ai Phasianidae dei Galliformi, è un genere raro ma importante endemico della Cina. Le quattro specie di Crossoptilon precedentemente riconosciute sono C. harmani , C. mantchuricum , C. crossoptilon e C. auritum. C. harmani è distribuito solo in habitat tipici alpini e di taiga ad altitudini di 3500–3900 metri nel Tibet sud-orientale. C. mantchuricum è distribuito principalmente nei monti Luliang della provincia dello Shanxi, nei monti Xiaowutai della provincia di Hebei, nei monti Dongling di Pechino e in alcune altre aree locali. C. crossoptilon è presente solo nella provincia occidentale del Sichuan, nella provincia nord-occidentale dello Yunnan, nella provincia sud-orientale del Qinghai e nel Tibet orientale in aree di alta montagna. È comune ad altitudini elevate (3000–4300 m) ed esiste nelle foreste di conifere, nelle foreste miste di latifoglie e conifere e nelle macchie alpine. C. auritum si incontra solo nelle regioni montuose delle province di Qinghai, Gansu e Sichuan e nella regione autonoma di Ningxia Hui.
Lo stato tassonomico e la posizione filogenetica di C. harmani sono controversi. Mentre alcuni studi suggeriscono che si tratti di una specie indipendente, altri propongono che si tratti di una sottospecie di C. crossoptilon. Si ritiene che il taxon C. mantchuricum si sia differenziato per primo, e C. crossoptilon e C. auritum sono considerati più strettamente correlati rispetto alle altre specie. Altri studi indicano che C. crossoptilon possa essere una specie relativamente originale e che C. auritum e C. mantchuricum siano più strettamente correlati geneticamente.
Studi precedenti sulle specie di Crossoptilon si sono concentrati sulla loro distribuzione, ecologia e comportamento. Per alcuni sono stati utilizzati singoli geni o sequenze parziali allo scopo di ricostruire le loro relazioni filogenetiche, ma raramente è stato tentato uno studio completo dell'intero mitogenoma. Per ricostruire le relazioni filogenetiche tra Crossoptilon e altre specie di Phasianidae, sono stati sequenziati tutti i mitogenomi di C. harmani , C. mantchuricum e C. crossoptilon, e quindi quattro specie del genere Crossoptilon sono state accuratamente confrontate. Inoltre, sono stati calcolati i tempi di divergenza stimati, ed è stata utilizzata un'analisi Ka/Ks per stimare l'adattamento del mitogenoma Crossoptilon a diversi ambienti.."
Metodi:
- Raccolta dei campioni ed estrazione del DNA
"..Campioni di C. harmani sono stati raccolti nell'area di Nagqu in Tibet, Cina, e un esemplare di prova è stato depositato presso Institute of Zoology, Shaanxi Normal University; C. mantchuricum e C. crossoptilon sono stati raccolti dallo Zoo di Pechino, e campioni sono stati ottenuti dalle collezioni di esemplari di uccelli del National Zoological Museum, Institute of Zoology, Chinese Academy of Sciences. La raccolta è avvenuta con il permesso del Dipartimento Forestale ed è stata conforme alla Legge Nazionale sulla Conservazione della Fauna Selvatica in Cina. Non sono stati condotti esperimenti su animali vivi nella presente ricerca. Tutti i campioni sono stati conservati in etanolo al 100% e preservati a -20°C. Il DNA genomico totale è stato estratto dal fegato/tessuto muscolare utilizzando il metodo standard fenolo/cloroformio.."
- Amplificazione e sequenziamento PCR
"..Le PCR sono state eseguite nelle seguenti condizioni: denaturazione iniziale di 2 minuti a 93°C, 40 cicli di: denaturazione di 10 s a 92°C, annealing di 30 s a 58–53°C, allungamento di 10 minuti a 68°C nei 20 cicli preliminari e denaturazione di 10 s a 92°C, annealing di 30 s a 53°C e allungamento di 10 minuti a 68°C con 20 s per ciclo aggiunti alla fase di allungamento nei successivi 20 cicli e infine un'estensione per 7 minuti a 68°C. Le amplificazioni sono state eseguite in reazioni da 15 μL contenenti 2,4 μL di dNTP 2,5 mM, 2,1 μL di ciascun primer a 10 μΜ, 1,5 μL di tampone PCR I 10× LA (senza Mg 2+), 1,5 μL di MgCl 2 25 mM , 1 μL di stampo di DNA, 0,18 μL di Taq polimerasi LA 5 U/μl (Takara, Dalian, Cina) e 4,22 μL di ddH 2 O.
Le sequenze dei primer utilizzati per l'amplificazione PCR e il sequenziamento dei geni mitocondriali sono state ottenute da Sorenson (2003) con piccole modifiche (file aggiuntivo 1 ). Il mitogenoma di C. harmani è stato amplificato in sette parti e le lacune sono state colmate utilizzando altri primer adiacenti. I prodotti della PCR sono stati purificati utilizzando il DNA Agarose Gel Extraction Kit (Bioteke, Pechino, Cina) dopo separazione mediante elettroforesi su un gel di agarosio allo 0,8%. Dopo la separazione e la purificazione, i prodotti della PCR sono stati sequenziati da Sangon Biotech (Shanghai) Co., LTD. utilizzando il metodo Primer-Walking. I mitogenomi di C. mantchuricum e C. crossoptilon sono stati amplificati in 12 o 13 frammenti e il sequenziamento è stato eseguito utilizzando il sistema di sequenziamento ad alto rendimento Illumina Hiseq2000 di Shenzhen Huada Gene Technology Co., LTD. Le lacune nell'assemblaggio dopo il sequenziamento ad alto rendimento sono state colmate mediante sequenziamento diretto utilizzando il sequenziatore di DNA ABI 3730 di Sangon Biotech (Shanghai) Co., LTD., utilizzando primer PCR adiacenti.."
- Identificazione genica e analisi del genoma
"..Il pacchetto di analisi della sequenza Staden è stato utilizzato per l'assemblaggio della sequenza e l'annotazione del mitogenoma di C. harmani. Gli assemblaggi genomici completi per i mitogenomi di C. mantchuricum e C. crossoptilon sono stati eseguiti utilizzando il software SOAP de novo. La maggior parte dei geni tRNA è stata identificata utilizzando tRNAscan-SE 1.21 nella modalità di ricerca 'solo tRNAscan', con il codice genetico mitocondriale dei vertebrati e la fonte 'mito/cloroplasto'. I geni codificanti proteine (PCG), i geni rRNA e i restanti geni putativi tRNA che non sono stati identificati da tRNAscan-SE sono stati identificati mediante confronto di sequenza con altre specie di Galliformes. La struttura secondaria rrnS di Crossoptilon è stata prevista sulla base della struttura di Gallus gallus e Anas platyrhynchos ottenuta dal Comparative RNA Web (CRW) e della struttura di Pseudopodoces humilis (ora come Parus humilis ). La struttura secondaria rrnL è stata prevista sulla base della struttura di Xenopus laevis ottenuta dal database CRW, delle strutture di Bos taurus e P. humilis. Il software RNAstructure è stato utilizzato per identificare e disegnare potenziali strutture secondarie nella regione di controllo a singolo filamento. Le composizioni nucleotidiche dei mitogenomi e le informazioni sugli amminoacidi sono state analizzate utilizzando MEGA 4.1.."
- Allineamenti di sequenza
"..Insieme all'intero mitogenoma ottenuto in questo studio, sono state utilizzate 42 sequenze di Galliformes nell'analisi filogenetica, inclusi due outgroup (Numida meleagris e Alectura lathami). Le sequenze di DNA delle altre specie utilizzate nelle analisi filogenetiche sono state scaricate da GenBank (i numeri di accesso e le informazioni chiave sono mostrati nel file aggiuntivo 2). I geni tRNA e rRNA e il CR sono stati allineati individualmente utilizzando ClustalX 1.83 con le impostazioni predefinite. Tutti i 13 geni codificanti proteine sono stati tradotti in amminoacidi e quindi allineati utilizzando MEGA 4.1 con parametri predefiniti per ciascun gene e infine ritradotti in sequenze nucleotidiche.."
- Analisi filogenetiche dei Phasianidae
"..I set di dati contenenti 13 geni codificanti proteine (PCG) e tutti i 37 geni più la regione di controllo (mitogenoma) sono stati utilizzati per studiare le relazioni filogenetiche all'interno dei Phasianidae. L'analisi filogenetica basata sulle sequenze nucleotidiche è stata eseguita utilizzando PAUP*4.0b10 per il metodo di massima parsimonia (MP), RAxML-7.0.3 per la massima verosimiglianza (ML) e MrBayes 3.1.2 per l'inferenza bayesiana (BI). Per le analisi ML e BI, i modelli dei set di dati delle sequenze nucleotidiche concatenate sono stati valutati indipendentemente utilizzando AICc in MrModeltest2.2. Il modello più adatto GTR + I + G è stato scelto per le analisi di verosimiglianza e bayesiane. Un albero di consenso è stato generato per l'analisi MP secondo la regola della maggioranza. L'affidabilità dei cladi negli alberi filogenetici è stata valutata mediante probabilità di bootstrap (BSP) calcolate utilizzando 1000 repliche, con aggiunta casuale per ciascuna replica di bootstrap. Il supporto bootstrap di 1000 repliche è stato eseguito anche nell'analisi ML. L'analisi bayesiana con campionamento Markov Chain Monte Carlo è stata eseguita per 1.000.000 di generazioni, salvando un albero ogni 100 generazioni, con una catena fredda e tre catene riscaldate, e il tempo di burn-in è stato determinato dal tempo di convergenza dei punteggi di verosimiglianza. Le probabilità posteriori bayesiane (BPP) sono state stimate su un albero di consenso con regola di maggioranza al 50% degli alberi rimanenti.
Abbiamo esaminato le prestazioni dei singoli geni e dei set di dati basati sui nucleotidi; le analisi PBS sono state eseguite nel programma che combina TreeRot.v3 e PAUP*4.0b10. I seguenti set di dati sono stati utilizzati per l'analisi: 13 geni codificanti proteine, rrnS , rrnL , partizioni CR, il primo, secondo e terzo codone di PCG, i tre cluster di geni tRNA (IQM, WANCY e HSL), ATP (atp6 + atp8), COX ( cox1 + cox2 + cox3 ) e NADH ( nad1 + nad2 + nad3 + nad4 + nad4L + nad5 + nad6 ).
Il MEGA 4.1 è stato utilizzato per calcolare la distanza genetica a coppie per quattro specie di Crossoptilon con parametri predefiniti. I dati allineati del mitogenoma e quattro singoli geni (nad2 , CR, cytb e rrnS) ottenuti da GenBank (file aggiuntivo 3 ) sono stati utilizzati per calcolare le distanze genetiche. Questi geni sono stati allineati singolarmente e possono essere regolati manualmente su lunghezze di sequenza coerenti.."
- Stime del tempo di divergenza focalizzate su Crossoptilon
"..Insieme al set di dati PCG ottenuto in questo studio, sono state utilizzate 42 sequenze di Galliformes per stimare il tempo di divergenza della specie Crossoptilon calcolato utilizzando la procedura bayesiana implementata in BEAST v. 1.7.2. È stato utilizzato un orologio rilassato con tassi conformi a una distribuzione log-normale. Il modello GTR + I + G e una priorità di Yule sono stati utilizzati nell'analisi. I punti di calibrazione erano basati sui registri fossili che mostravano che lo stelo Numididae-Phasianidae si è diviso a 50-54 Mya (milioni di anni fa); Arborophila rufipectus diverge dagli altri lignaggi nei Galliformes intorno a 39 Mya; Coturnix - Gallus si è diviso a 35 Mya. I risultati di serie di 10 milioni di generazioni sono stati utilizzati dopo un burn-in di 100.."
- Analisi Ka e Ks
"..Per comprendere meglio l'evoluzione a livello del DNA e il ruolo della selezione nelle quattro specie di Crossoptilon, abbiamo calcolato i tassi di sostituzione non sinonimi e sinonimi usando Kaks_calculator 2.0 per sei gruppi [ C. harmani - C. mantchuricum ( C.har - C.man ), C. mantchuricum - C. crossoptilon ( C.man -C.cro ), C. harman i- C. crossoptilon ( C.har - C.cro ), C. harmani - C. auritum ( C.har - C.aur ), C. mantchuricum - C. auritum ( C.man - C.aur ), e C. crossoptilon - C. auritum ( C.cro - C.aur )]. Il rapporto tra il tasso di sostituzione non sinonimico (Ka) e il tasso di sostituzione sinonimo (Ks) è ampiamente utilizzato come indicatore della pressione selettiva a livello di sequenza tra specie diverse. È comunemente accettato che Ka > Ks, Ka = Ks e Ka < Ks indichino generalmente selezione positiva, mutazione neutra e selezione negativa, rispettivamente. Per calcolare Ka, Ks e Ka/Ks, è stato selezionato un metodo di media del modello. Questo metodo include 14 diversi modelli per il calcolo e ha derivato i valori medi per Ka, Ks e Ka/Ks. Il codice genetico selezionato è stato il "codice mitocondriale dei vertebrati". Per studiare ulteriormente la pressione selettiva agita su ciascun gene codificante per proteine nel genere Crossoptilon, è stato utilizzato CodeML nel software PAMLX per trovare siti sotto forte pressione selettiva. L'analisi della struttura secondaria dell'amminoacido è stata eseguita utilizzando un software online TOPCONS.."
- Risultati
"..Il confronto tra i due metodi di sequenziamento utilizzati in questo studio ha rivelato che il sequenziamento ad alto rendimento (HTP) offre una copertura e un'accuratezza maggiori rispetto al sequenziamento standard, sebbene a un costo maggiore. Utilizzando il sequenziamento standard, abbiamo ottenuto 62 sequenze efficaci con una copertura di 2,63 volte superiore. Al contrario, il sequenziamento ad alto rendimento (HTP) ha prodotto dati di assemblaggio efficaci con profondità di sequenza (X) di 7604,35 per C. mantchuricum e 7810,82 per C. crossoptilon, dopo aver filtrato alcune letture, come quelle di bassa qualità o contaminate da sequenze adattatrici.
I mitogenomi completi di C. harmani , C. mantchuricum e C. crossoptilon sono rispettivamente lunghi 16682 bp, 16690 bp e 16680 bp. I genomi annotati sono stati depositati nel database GenBank (numeri di accesso: KP259806-KP259808 ). I geni mitocondriali di tre specie di Crossoptilon sono codificati sul filamento H, ad eccezione di un gene codificante per proteine (nad6) e otto geni tRNA ( trnQ , trnA , trnN , trnC , trnY , trnS (UCN) , trnP e trnE ) (Tabella 1 ). In linea con i risultati precedenti sui mitogenomi dei Phasianidae, le sequenze mitocondriali di quattro specie di Crossoptilon ( C. harmani , C. mantchuricum , C. crossoptilon e C. auritum ) sono sbilanciate verso adenina e timina (rispettivamente 54,3%, 54,2%, 54,3% e 54,2%). L'asimmetria GC nei mitogenomi di Crossoptilon è simile, con parti di CR e rrnS che contengono asimmetrie significativamente più elevate rispetto ad altre regioni. Le regioni CR, trnF , rrnS , trnV , rrnL e cox1 presentano forti asimmetrie GC.."
- Geni che codificano proteine
"..I 13 geni codificanti proteine dei genomi di Crossoptilon sono simili a quelli della maggior parte delle altre specie di Phasianidae, con nad5 e atp8 rispettivamente come geni più lunghi e più corti. La lunghezza totale dei PCG in ciascuna specie di Crossoptilon è di 11358 bp dopo la rimozione dei codoni di terminazione, contenenti circa 3786 codoni. Il contenuto di A+T dei 13 PCG è del 53,4% in C. harmani e C. crossoptilon, e del 53,3% in C. auritum e C. mantchuricum. L'analisi della composizione in basi in ciascuna posizione dei codoni dei PCG concatenati mostra che la seconda posizione del codone ha un contenuto di A+T più elevato (rispettivamente 57,9% in C. auritum e C. mantchuricum , 58,1% in C. harmani e 58,0% in C. crossoptilon ) rispetto alla prima e alla terza posizione del codone. Le frequenze degli amminoacidi nei PCG di Crossoptilon sono simili, con Leu significativamente più frequente rispetto agli altri amminoacidi. Le diverse posizioni dei codoni hanno la stessa distribuzione di basi, con C e G rispettivamente come basi più e meno frequenti nel terzo codone.
Tutti i PCG nelle specie di Crossoptilon iniziano con il tipico codone ATG, ad eccezione del gene nad5 in C. mantchuricum e del gene cox1 in quattro specie di Crossoptilon, che iniziano con GTG. I geni codificanti utilizzano quattro tipi di codoni di stop, tra cui TAA e TAG per la maggior parte dei geni, AGG per cox1 in Crossoptilon e nad6 in C. harmani e C. crossoptilon, e un codone di stop incompleto T- per cox3 , nad4 e nad2 in quattro specie di Crossoptilon, rispettivamente.."
- Geni dell'RNA
"..Analogamente ai mitogenomi precedentemente sequenziati, i genomi sequenziati in questo studio contengono 2 geni rRNA che codificano le subunità piccola e grande dell'rRNA, situate tra trnF e trnL(UUR) e separate dal gene trnV . Le lunghezze e il contenuto di A+T di rrnS e rrnL nei genomi di Crossoptilon rientrano nell'intervallo osservato in altre specie di Phasianidae. rrnS contiene tre domini con 46 staminali previste, mentre rrnL contiene sei domini con 59 staminali (file aggiuntivi 4 e 5 ). Le strutture secondarie di rrnS in C. crossoptilon e C. harmani sono identiche; le strutture secondarie di rrnL differiscono di sole 2 bp in lunghezza. Tuttavia, ci sono molte differenze nelle strutture secondarie di rrnL tra le quattro specie di Crossoptilon, in particolare nell'ansa vicino allo stelo 44, che contiene diverse sostituzioni e indel.
La sequenza completa del mitogenoma contiene 22 geni di tRNA intervallati. Tutte le sequenze di tRNA hanno il potenziale per ripiegarsi in tipiche strutture secondarie a trifoglio, ad eccezione di trnS(AGY) , che è privo del braccio DHU (file aggiuntivo 6 ). Le strutture secondarie di trnF , trnL(UUR) , trnQ , trnM , trnC , trnY , trnS(UCN) , trnH , trnL(CUN) , trnT e trnP sono relativamente conservate. Tuttavia, la struttura di trnS(AGY) è diversa in C. auritum rispetto alle altre tre specie di Crossoptilon (file aggiuntivo 6 ); la struttura in C. harmani , C. mantchuricum e C. crossoptilon contiene due basi aggiuntive (G e A) nel braccio accettore degli amminoacidi. La discrepanza più frequente è GU; sono presenti anche altre discrepanze, tra cui UU in trnM e trnG , CC in trnD e trnL(UUR) , CU in trnF e trnI e AC in trnS(AGY) . Le strutture dei tRNA di C. crossoptilon e C. harmani sono quasi identiche, ad eccezione di 1 bp. Il contenuto di A+T dei geni dei tRNA è del 57,6% in C. harmani e C. crossoptilon , del 57,4% in C. mantchuricum e C. auritum.."
Regione di controllo
"..La composizione nucleotidica della regione di controllo nella specie Crossoptilon ha una propensione contro G, che è comune nel filamento senso del mitogenoma dei vertebrati. La regione di controllo di Crossoptilon si trova nella posizione conservata tra trnE e trnF e la lunghezza (1146 nucleotidi) è conservata in tutte e quattro le specie. La regione di controllo contiene tre domini: il dominio ETAS I (nt 1–312), il dominio centrale conservato II (nt 313–780) e il dominio CSB III (nt 781–1146). Il contenuto di A + T dei tre domini è simile in tutte le specie di Crossoptilon; i domini I, II e III hanno contenuti più elevati di C e A, T e A, rispettivamente, rispetto ad altre basi. Il dominio III contiene una percentuale più alta di A rispetto al dominio II e il dominio II ha il contenuto di G più alto tra i tre domini. La distribuzione dei siti variabili e dei siti conservati suggerisce che il Dominio II ha relativamente più siti conservati e il Dominio I ha più siti variabili rispetto agli altri domini.
Il dominio I di ETAS può essere diviso in due parti: parte A, da nt 1–163 e parte B, da nt 164–312. Ci sono due blocchi conservati nella parte A, ETAS1 (nt 64–126) ed ETAS2 (nt 124–163), che sono simili a motivi precedentemente identificati in altre specie aviarie e di mammiferi. Il primo blocco è perfettamente conservato tra le specie di Crossoptilon e ha una similarità di sequenza con la “forcina d'oca” descritta in alcuni fasianidi e specie di Anas (file aggiuntivo 7 ). La struttura secondaria di questa forcina è determinata da uno stelo di sette Cs/G complementari e da un'ansa contenente un motivo TCCC presente anche nella regione di controllo dei mammiferi, che è stata associata sperimentalmente alla terminazione dei filamenti H. Ci sono due copie di TCCC situate nei nt 22–25 e 183–186. Il secondo blocco, che è perfettamente conservato tra le specie di Crossoptilon, ha una similarità di sequenza con i TAS dei mammiferi, incluso il motivo altamente conservato GTGCAT, che è presente in tutti i Fasianidi e Anseriformi sequenziati. Il motivo GYRCAT (Y = C/T; R = A/G) è diffuso nel Dominio I di alcune regioni di controllo dei mammiferi, ed è stato duplicato nelle ripetizioni R1 di molte specie, tra cui l'opossum e diversi roditori. La sua importanza funzionale è suggerita sia da dati comparativi che sperimentali. Il Dominio I a singolo filamento può formare potenziali strutture secondarie a trifoglio. I primi 190 nucleotidi del Dominio I delle specie di Crossoptilon possono formare strutture secondarie stabili (file aggiuntivo 7 ). Le configurazioni dei trifogli sono simili in C. harmani e C. crossoptilon e variano leggermente in C. mantchuricum e C. auritum; le strutture a stelo-ansa dei trifogli includono sempre la "forcina d'oca". La regione di controllo di C. harmani e C. crossoptilon presenta strutture simili con cinque steli, mentre C. mantchuricum e C. auritum ne hanno solo quattro.
Il dominio centrale conservato II della regione di controllo della specie Crossoptilon include 468 nucleotidi conservati, dal sito ipervariabile in posizione 313 alla fine del dominio ETAS all'inizio del presunto blocco di sequenza O H in posizione 781. Diversi blocchi conservati del dominio centrale conservato sono simili alle caselle F (nt 350–377), E (nt 395–414), D (nt 455–479) e C (nt 504–529) dei vertebrati e di altre specie aviarie, ed è presente anche una casella di similarità degli uccelli.
Il dominio III del CSB è altamente variabile e presenta sequenze simili a quelle del CSB1 dei mammiferi. Una sequenza poli(C) (nt 781–792), simile all'OH dei mammiferi, mappa solo pochi nucleotidi a valle del presunto CSB1 (nt 803–828). Tuttavia, è difficile identificare sequenze corrispondenti a CSB2 e CSB3 dei mammiferi. Le strutture secondarie di CSB1 e della presunta forcina d'oca in Crossoptilon sono coerenti con quelle di G. gallus (file aggiuntivo 7 ). I promotori bidirezionali LSP/HSP (nt 982–1003) sono quasi perfettamente conservati tra le specie di Crossoptilon. Una forcina stabile (nt 1004–1017) ricca di stringhe poli(T) e poli(A) si trova immediatamente a monte dei promotori.."
Filogenesi e tempo di divergenza della specie Crossoptilon
"..Il dataset PCG combinato finale ha 11376 caratteri dopo l'allineamento. Per questo dataset, l'analisi di parsimonia mostra una lunghezza di 27412 passaggi, con CI = 0,313, RI = 0,499. Le topologie tra gli alberi ML e BI del dataset PCG erano coerenti (Figura 1 ). Per il dataset del mitogenoma, non c'era differenza nella topologia tra gli alberi ML e BI di Galliformes (file aggiuntivo 8 ); tuttavia, entrambi gli alberi differivano dall'albero MP (file aggiuntivo 8 ). Secondo i risultati filogenetici, la monofilia di Crossoptilon è stata fortemente supportata nelle analisi MP, ML e BI. All'interno di Phasianidae, la topologia (( Crossoptilon + Lophura ) + ( Phasianus + Chrysolophus )) è stata formata nella maggior parte degli alberi. La relazione di gruppo fratello tra Crossoptilon e Lophura è stata supportata da valori bootstrap pari a 69 e 100 negli alberi MP e ML e da probabilità posteriori pari a 1,00 nell'albero BI del dataset PCG.


Le distanze genetiche dei mitogenomi (file aggiuntivo 9 ) sono identiche (0,026) per C. auritum e C. harmani , C. auritum e C. crossoptilon , C. harmani e C. mantchuricum e C. mantchuricum e C. crossoptilon. Tuttavia, le distanze genetiche tra C. harmani e C. crossoptilon e C. auritum e C. mantchuricum sono rispettivamente solo 0,001 e 0,002. Le distanze genetiche basate su quattro singoli geni mostrano che le distanze genetiche di C. harmani - C. crossoptilon e C. auritum - C. mantchuricum sono molto più piccole. Questo valore è simile al livello intraspecifico e molto inferiore al livello interspecifico (file aggiuntivo 9 ).
Secondo la scala temporale stimata ottenuta dall'albero filogenetico contenente una topologia coerente con gli alberi PCG-ML/BI, le separazioni C. harmani - C. crossoptilon e C. mantchuricum - C. auritum si sono verificate a 3,21 milioni di anni fa (95% della più alta densità di probabilità a posteriori (HPD) = 2,36-4,23 milioni di anni fa) (Figura 2 ). Il tempo di divergenza per C. harmani e C. crossoptilon è di circa 0,11 milioni di anni fa (95% HPD = 0,05-0,18 milioni di anni fa), per C. mantchuricum e C. auritum, 0,18 milioni di anni fa (95% HPD = 0,1-0,29 milioni di anni fa) (Figura 2 ).."
- Sostituzione sinonima e non sinonima
"..L'analisi dei siti variabili in ciascun gene codificante per proteine ha mostrato che il gruppo C.har - C.cro e C.man - C.aur conteneva meno siti variabili, mentre altri quattro gruppi ne includevano di più (file aggiuntivo 10 ). Le percentuali di siti variabili erano più alte nei geni nad3 e nad6 tra i gruppi C.har - C.man , C.man - C.cro , C.har - C.aur e C.cro - C.aur , mentre le percentuali erano più basse nei geni atp8 e atp6 . Il valore Ka/Ks di atp8 nei cinque gruppi è di gran lunga maggiore di 1, eccetto che per C.har - C.cro , e atp6 e cytb in C.har - C.cro (Tabella 2 - non allegata), che mostrano una forte selezione positiva. I valori Ka/Ks degli altri geni analizzati nelle specie Crossoptilon erano inferiori a 1, il che mostra una selezione purificante (Tabella 2 - non allegata). Inoltre, il valore P (test esatto di Fisher) è molto inferiore a 0,001, eccetto per atp8 in C.har - C.man , C.man - C.cro , C.har - C.aur e C.cro - C.aur (rispettivamente 0,558274 o 0,289204, che sono ovviamente superiori a 0,05) e nad3 in C.har - C.cro (0,154), indicando che la differenza è significativa.."
"..Entrambi i geni atp6 e cytb hanno una base diversa nel gruppo C.har - C.cro , ovvero nt 245 nel gene atp6 (base T o Phe in C. harmani, mentre base C o Ser in C. crossoptilon) e nt 709 nel gene cytb (base C o Leu in C. harmani, mentre base T o Phe in C. crossoptilon). Anche il gene atp8 nel gruppo C.man - C.aur contiene una base diversa (nt 124), base G in C. mantchuricum, mentre base A in C. auritum, e di conseguenza Val in C. mantchuricum, mentre base Met in C. auritum. Ulteriori studi hanno dimostrato che quattro siti amminoacidici in atp8 (T in posizione 11, I in posizione 12, S in posizione 39 e V in posizione 42) nel genere Crossoptilon erano siti selezionati positivamente, sulla base dell'analisi Bayes Empirical Bayes (BEB). Questi quattro diversi siti amminoacidici (T, I, S e V) corrispondevano alle sequenze nucleotidiche di ACT, ATC, AGC e GTA in C. harmani e C. crossoptilon, ATT, ACC, AAC e GTA in C. mantchuricum, ATT, ACC, AAC e ATA in C. auritum. L'ulteriore analisi della struttura secondaria della sequenza amminoacidica di atp8 ha mostrato che T in posizione 11 e I in posizione 12 si trovano nel dominio elica (IN->OUT) nella proteina transmembrana (TM), mentre S in posizione 39 e V in posizione 42 si trovano nel dominio esterno.."
Analisi PBS
"..Analisi PBS sono state eseguite per comprendere meglio il contributo delle diverse parti dei mitogenomi alla filogenesi del genoma basata sull'albero mitogenoma-ML. Sono state studiate anche le relazioni tra valore PBS, lunghezza, siti singleton (S), siti informativi di parsimonia (Pi), siti variabili (V) e siti conservati (C) in diverse partizioni. La classificazione dei singoli geni codificanti proteine in base al rispettivo contributo ai valori PBS totali mostra che alcuni geni, come nad5 e nad4, forniscono un contributo maggiore rispetto ad altri marcatori, mentre atp8 e nad4L contribuiscono meno. Tuttavia, l'analisi mostra anche che i valori PBS sono approssimativamente correlati con la lunghezza dei geni. I siti variabili sono strettamente correlati ai siti informativi di parsimonia, che seguono la stessa tendenza. Il terzo codone ha un valore PBS elevato ma con meno siti conservati, mentre il cluster genico IQM dei tRNA ha un valore PBS inferiore con più siti conservati rispetto ad altre partizioni.
Il terzo codone ha un valore PBS elevato e quindi contribuisce maggiormente all'albero mitogenomico-ML. I geni NADH presentano un valore PBS secondario elevato, che potrebbe essere l'effetto del terzo codone nella sequenza relativamente lunga. Per comprendere meglio il contributo del terzo codone, abbiamo ricostruito l'albero filogenetico utilizzando questa singola partizione.
Negli alberi filogenetici basati sul terzo codone (file aggiuntivo 8), la struttura dell'albero ML è simile a quella del BI, ad eccezione di Phasianus. Le posizioni di Pucrasia macrolopha, Bonasa bonasia, Meleagris gallopavo e Polyplectron bicalcaratum nel terzo albero MP sono diverse rispetto all'albero BI. Il terzo albero BI è identico al mitogenome-ML con un supporto relativamente elevato, ad eccezione della posizione di Polyplectron bicalcaratum.."
Discussione
Relazione filogenetica e tempo di divergenza della specie Crossoptilon
"..In questo studio, gli alberi filogenetici basati su diversi set di dati supportano la monofilia di Crossoptilon e le strette relazioni genetiche di C. auritum e C. mantchuricum, C. harmani e C. crossoptilon. La nostra analisi delle sequenze geniche di Crossoptilon basata su diversi set di dati conferma che Crossoptilon è il gruppo fratello di Lophura, indicando la loro relazione più stretta.
La distanza genetica è piuttosto piccola tra alcune specie di Crossoptilon, ad esempio, tra C. harmani e C. crossoptilon , C. auritum e C. mantchuricum. Studi morfologici, ecologici e comportamentali hanno rivelato l'elevata somiglianza tra C. harmani e C. crossoptilon, ma con diversa colorazione del piumaggio. Le loro distribuzioni sono sovrapposte e C. harmani si ibrida regolarmente con C. c. drouynii, il che indica la loro compatibilità riproduttiva. Inoltre, la distribuzione di C. harmani è limitata tra le catene himalayane e Nyenchen Tanglha, che hanno fornito l'isolamento geografico richiesto per la formazione di una sottospecie o specie. Pertanto, lo stato tassonomico e la sua relazione con C. crossoptilon sono molto discutibili e devono essere rivalutati da più marcatori e dalla genetica di popolazione negli studi successivi.
Sulla base delle loro significative differenze in molti aspetti, come la morfologia e il comportamento, C. auritum e C. mantchuricum sono comunemente considerate specie indipendenti. Tuttavia, la distanza genetica di 0,002 indica che hanno una relazione genetica molto più stretta. In linea con questa osservazione, Tsam et al. (2003) hanno osservato che la differenziazione interspecifica tra C. mantchuricum e C. auritum (0,18%) è inferiore al grado di differenziazione tra le sottospecie di C. crossoptilon.
Alcune specie di Phasianidae sono state trovate nell'epoca del Pliocene e diversi taxa moderni erano già apparsi nel Pleistocene quaternario. I primi fossili di Crossoptilon sono stati trovati negli strati del Cenozoico Zhoukoudian di Pechino e negli strati del Pleistocene a Yanjinggou nel Wanxian della provincia del Sichuan. L'evoluzione di Crossoptilon si è verificata principalmente in corrispondenza dell'intenso sollevamento dell'altopiano tibetano durante il Pliocene terziario e dell'alternanza dei periodi glaciali e interglaciali nel Quaternario, che ha creato profondi e complessi cambiamenti negli ambienti geografici ed ecologici e si pensa che abbia influenzato notevolmente la sua topografia così come la diversificazione delle specie aviarie. Sulla base dei nostri risultati, il tempo di separazione tra C. harmani - C. crossoptilon e C. mantchuricum - C. auritum è coerente con il sollevamento durante il Pliocene terziario e con il record fossile negli strati del Pleistocene Yanjinggou Wanxian del Sichuan, che è approssimativo ai tempi di divergenza stimati da Jiang et al. (2014) (3,78 Mya, 95% HPD = 1,17-6,56 Mya). Il nostro studio mostra che C. mantchuricum e C. auritum sono gruppi relativamente antichi, mentre C. harmani e C. crossoptilon hanno divergenza più tardi; queste osservazioni non sono coerenti con l' ipotesi evolutiva di Crossoptilon proposta da Lu et al. (1998) .
Lu et al. (1998) suggeriscono che l'antenato di C. crossoptilon e C. harmani abbia avuto origine nelle provincie del Sichuan, Yunnan e nell'area di confine tibetana. Notarono che il clade contenente la plesiomorfia di C. harmani è migrato verso l'altopiano dell'entroterra e successivamente si è differenziato nell'antenato di C. harmani e C. auritum. L'antenato del C. auritum si è disperso nell'altopiano settentrionale e si è differenziato in C. auritum e C. mantchuricum. Sulla base del tempo di divergenza e delle caratteristiche di distribuzione delle specie di Crossoptilon, i nostri risultati supportano che l'antenato di Crossoptilon fosse inizialmente distribuito nelle provincie del Sichuan, Yunnan e l'area di confine tibetana. Il Crossoptilon si è inizialmente diversificato nell'antenato di C. harmani - C. crossoptilon e nell'antenato di C. auritum - C. mantchuricum. Considerando la distribuzione geografica di Crossoptilon, proponiamo che l'antenato di C. harmani - C. crossoptilon potrebbe essere migrato sulla piattaforma dell'altopiano e poi essersi diviso in due lignaggi durante l'ultimo sollevamento del Plateau tibetano, uno adattato all'ambiente di alta quota per formare C. harmani nel Tibet orientale, l'altro evolutosi in C. crossoptilon, mentre l'antenato di C. auritum - C. mantchuricum si è disperso sull'altopiano settentrionale differenziandosi ulteriormente quasi nello stesso periodo con C. harmani - C. crossoptilon.."
- Pressione selettiva sui geni codificanti proteine nelle specie Crossoptilon
"..I valori di Ka/Ks nella maggior parte dei geni codificanti proteine sono inferiori a 1 (Ka è inferiore a Ks) con valori di P inferiori a 0,001, il che indica che questi geni in Crossoptilon sono sottoposti a selezione purificante. I valori di Ka/Ks di ciascun gene tra C.har - C.man , C.man - C.cro , C.har - C.aur e C.cro - C.aur sono simili. Il valore di Ka/Ks del gene atp8 è 50 in cinque gruppi, ad eccezione di C.har - C.cro , il che rivela una forte selezione positiva. Tuttavia, solo il valore di P per C.man -C.aur è molto inferiore a 0,01 (P = 0), il che indica differenze significative. Al contrario, i valori di Ka/Ks dei geni atp6 e cytb in C.har - C.cro sono 50 con valori di P inferiori a 0,001, il che indica anch'esso una forte selezione positiva. Studi precedenti hanno dimostrato che il gene atp6 è altamente variabile nei mitogenomi degli esseri umani che vivono in aree estremamente fredde, il gene atp6 affronta una forte pressione di selezione con l'aumento dell'altitudine e la variazione del gene atp6 in Artemia tibetiana è il risultato dell'adattamento all'ambiente freddo e ipossico dell'altopiano. La variazione in atp6 può riflettere l'adattamento di C. harmani all'ambiente dell'altopiano; cytb potrebbe anche svolgere un ruolo importante in questo processo. Anche il gene atp8 in C. mantchuricum e C. auritum potrebbe aver subito una forte selezione nell'adattamento all'ambiente dell'altopiano.
Tuttavia, lo studio successivo ha mostrato che solo quattro siti amminoacidici di atp8 nel Crossoptilon sono siti selezionati positivamente e, di conseguenza, questo gene potrebbe aver subito una forte pressione selettiva per l'adattamento al plateau. La corrispondente elica TM (IN-> OUT) o la posizione esterna potrebbero essere importanti per l' adattamento del Crossoptilon . Dopo aver scartato i gruppi ( C.har - C.cro e C.man - C.aur ) e i geni ( cox1 , atp8 e nad4l ) con valori Ka/Ks ovviamente diversi dagli altri, il valore medio (0,0431) è simile al gruppo fortemente locomotore (0,04) nello studio di Shen et al. (2009), il che indica che il Crossoptilon accumula meno mutazioni non sinonime e corrisponde alla loro forte capacità di corsa.."
- Conclusioni
"..In sintesi, i mitogenomi della specie Crossoptilon presentano la stessa disposizione genica e composizioni simili, inclusi il contenuto di basi e le strutture secondarie. Secondo gli alberi filogenetici, C. auritum con C. mantchuricum e C. harmani con C. crossoptilon formano due coppie di gruppi fratelli. Crossoptilon ha una relazione più stretta con Lophura. In base alle distanze genetiche, C. harmani è più strettamente correlato a C. crossoptilon ed è il discendente divergente più recente di Crossoptilon, a seguito dell'adattamento all'altopiano. Secondo i risultati della datazione molecolare, il tempo di divergenza tra C. harmani - C. crossoptilon e C. mantchuricum - C. auritum è coerente con il sollevamento dell'altopiano tibetano e il successivo cambiamento climatico durante il Pliocene Terziario. L'analisi Ka/Ks ha mostrato che il gene atp8 nella specie Crossoptilon potrebbe aver subito una forte selezione per l'adattamento all'ambiente dell'altopiano.."
Riassumendo ( studi del 2000 e 2015 ):
A) Crossoptilon auritum: il fagiano orecchiuto blu fu il primo del suo genere ad essere descritto ma, curiosamente, è sempre rimasto il meno conosciuto tra tutti i fagiani orecchiuti. Nel 1811 Pallas lo descrisse attribuendogli il nome di Phasianus auritus nella sua "Zoografia Rosso (Russo)-Asiatica". Il tipo è andato ormai perduto da tempo e la descrizione originale era così poco dettagliata che, quando furono scoperti esemplari della forma bruna (Crossoptilon mantchuricum), le due specie furono confuse per molti anni. Tuttavia, quando l'Abate David inviò due pelli di orecchiuto blu (Crossoptilons auritum) al Museo di Parigi, ogni dubbio fu dissipato sulla relativa distinzione delle due forme.
B) Crossoptilon crossoptilon: Hodgson, nel 1838, descrisse un fagiano bianco con il nome di Phasianus tibetanus. Ottenne questo uccello in Nepal da un inviato indigeno appena tornato da Pechino e poiché per molti anni questo esemplare fu l'unico, i suoi habitat rimasero per un periodo di tempo altrettanto lungo completamente avvolti nel mistero. Alla fine si scoprì che il tipico tibetanus non presentava macchie bianche sulle penne della coda, e quindi l'uccello di Hodgson non era assolutamente purosangue, ma presentava una certa traccia di auritum 1*).
C) Crossoptilon mantchuricum: venne descritto quasi per caso da Swinhoe nel 1962 che inizialmente riteneva l'esemplare esaminato un femmina di Crossoptilon auritum.
Commento personale:
1*) Potrebbe anche essere, considerato che crossoptilon e auritum si incontrano nel nord del Sichuan, tuttavia le "macchie" bianche sulla coda ed il piumaggio arruffato suggeriscono che si potesse trattare di un esemplare chiaro di dolani. Vedere descrizione originale nella parte dedicata al Crossoptilon c. crossoptilon.
a) Nello studio del 2015 lo status del Crossoptilon harmani è, nuovamente, messo in dubbio.
b) Oggi sappiamo che gli incroci, e probabilmente i re-incroci tra ibridi o tra quest'ultimi e le forme genitoriali, producono ibridi fecondi che mostrano le più disparate colorazioni dei piumaggi. Ciò suggerisce che il Crossoptilon c. dolani, o meglio, la forma che parrebbe rappresentarlo attualmente, appartenga ad uno di questi.
c) Non solo qualche esemplare, ma si possono facilmente osservare popolazioni più o meno numerose con esemplari dalla coda pressoché solo bianca ed un fenotipo in generale molto uniforme. Vedere immagini nella parte dedicata ad ibridi e forme incerte.
Segue il sunto di un altro interessante studio riguardante il Crossoptilon mantchuricum e auritum condotto nel 2021.
Comparative Study - Mol Biol Evol. 2021 Jan 4;38(1):263-273. doi: 10.1093/molbev/msaa213.
Genomic Consequences of Long-Term Population Decline in Brown Eared Pheasant
Pengcheng Wang, John T Burley , Yang Liu , Jiang Chang , De Chen , Qi Lu , Shou-Hsien Li , Xuming Zhou , Scott Edwards, Zhengwang Zhang.
PMID: 32853368 - PMCID: PMC7783171 - DOI: 10.1093/molbev/msaa213
Testo per esteso, figure, set di dati supplementari e tutto quanto non pubblicato per ragioni di spazio su questa pagina, sono consultabili al seguente link: https://pmc.ncbi.nlm.nih.gov/articles/PMC7783171/
Astratto "..La teoria genetica di popolazione e le prove empiriche indicano che gli alleli deleteri possono essere eliminati in piccole popolazioni. Tuttavia, questo punto di vista rimane controverso. Non è chiaro se la selezione naturale sia abbastanza potente da escludere le mutazioni deleterie quando le popolazioni selvatiche continuano a diminuire. I fagiani sono uccelli terrestri che affrontano un rischio di estinzione a lungo termine a causa di perturbazioni e sfruttamento antropici. Ciononostante, le risorse genomiche disponibili per la gestione e la pianificazione della conservazione sono scarse. In questo articolo, abbiamo analizzato i dati genomici comparativi di popolazione per le tre popolazioni isolate esistenti di fagiano orecchiuto bruno (Crossoptilon mantchuricum) in Cina. Abbiamo dimostrato che questa forma mostra una bassa diversità genomica ed una dimensione effettiva della popolazione in contrazione a causa dei persistenti cali negli ultimi 100.000 anni. Abbiamo confrontato la variazione genomica di C. mantchuricum con quella della sua specie sorella strettamente correlata, il fagiano orecchiuto blu (Crossoptilon auritum), per la quale la preoccupazione per la conservazione è bassa. Sono state riscontrate conseguenze genetiche dannose in tutti i genomi di Crossoptilon mantchuricum, tra cui sequenze omozigoti estese, bassi tassi di decadimento da disequilibrio di linkage, eccessive mutazioni con perdita di funzione e perdita di diversità genetica adattativa a livello della regione del complesso maggiore di istocompatibilità. Per quanto a nostra conoscenza, questo studio è il primo a condurre un'analisi genomica completa per la conservazione di questa specie di fagiano minacciata. Inoltre, abbiamo dimostrato che la selezione naturale potrebbe non essere sufficiente a eliminare le mutazioni deleterie nelle popolazioni selvatiche in declino a lungo termine. I risultati di questo lavoro potrebbero facilitare la pianificazione della conservazione delle specie minacciate e contribuire al recupero delle dimensioni della loro popolazione.."
Crossoptilon mantchuricum.- Tempio di Xuanzhong, Monti Lüliang Shanxi 30-08-2020 - www.shanghaibirding com Autore: Craig Brelsford

Crossoptilon auritum.- Contea di Luqu, prefettura autonoma tibetana del Gannan, Gansu 03-05-2023 Cina - www.inaturalist.org Autore Bīng Niǎo

Introduzione
"..La diversità genetica è un fattore chiave nella pianificazione delle misure di conservazione per le specie minacciate in quanto fornisce informazioni vitali per gli sforzi di conservazione (Frankham et al. 2014 ; Xue et al. 2015 ; Feng et al. 2019). Poiché i colli di bottiglia demografici portano a livelli più elevati di consanguineità e deriva genetica, ci si aspetta che una contrazione della popolazione possa comportare l'accumulo o persino la fissazione di mutazioni deleterie (Xue et al. 2015 ; Feng et al. 2019). Prove recenti hanno dimostrato che le specie minacciate con piccole dimensioni effettive della popolazione e bassa diversità genetica possono comunque persistere per migliaia di anni dopo un collo di bottiglia demografico (Robinson et al. 2016). La selezione purificante può esercitare un'influenza relativamente maggiore sulla rimozione degli alleli deleteri in popolazioni piccole e stabili (Robinson et al. 2016; Grossen et al. 2020 ). Tuttavia, non è certo se la deriva genetica riduca l'efficacia della selezione purificata nell'attenuare le conseguenze genetiche dannose che si verificano quando le popolazioni selvatiche subiscono un continuo declino.
I fagiani (Phasianidae; Galliformes) sono uccelli terrestri ampiamente distribuiti (Johnsgard 1986). Hanno svolto, e svolgono ruoli importanti nell'economia, cultura ed ecologia di varie società umane (Peng et al. 2016 ; Peters et al. 2016 ; Barton et al. 2020). Poiché il loro piumaggio è luminoso, la loro carne appetibile e la loro mobilità limitata, i fagiani sono stati ampiamente cacciati e sfruttati dall'uomo (Fuller e Garson 2000 ; Barton et al. 2020 ). Di conseguenza, alcune specie sono diminuite o estinte del tutto (Keane et al. 2005 ; McGowan et al. 2012). Come ad esempio, la quaglia neozelandese ( Coturnix novaezelandiae ) estintasi rapidamente entro il 1875 (BirdLife International 2016a).
Tuttavia, sono pochi gli studi pubblicati che documentano le conseguenze genomiche delle fluttuazioni demografiche storiche e della recente attività antropica sulle popolazioni di fagiani minacciate.
Il fagiano orecchiuto bruno (Crossoptilon mantchuricum) è una specie vulnerabile e in declino (BirdLife International 2016c). Compare nella Lista Rossa IUCN del 2020 delle specie minacciate ed è confinato a tre aree frammentate nelle province di Shaanxi, Shanxi e Hebei e nella municipalità di Pechino, nella Cina centro-settentrionale (fig. 1 a) ( Zheng 2015 ). Queste aree hanno subito continue perturbazioni antropiche negli ultimi 40.000 anni (40 ka) (Shang et al. 2007 ; Barton et al. 2020). Le capitali delle dinastie Qin (221-207 a.C.), Ming e Qing (1368-1912 d.C.) erano situate nello Shaanxi e Pechino. Durante questi periodi si verificò una massiccia deforestazione per consentire la costruzione di palazzi e case. I Viaggi di Marco Polo e altre fonti letterarie documentano l'uso delle penne caudali di C. mantchuricum per decorare i cappelli ufficiali dei generali. Questo lungo periodo di sfruttamento e distruzione dell'habitat potrebbe aver limitato la distribuzione di C. mantchuricum e causato la sua bassa diversità genetica nucleare e mitocondriale (Wang et al. 2017). Pertanto, la lunga storia di minacce nei confronti del C. mantchuricum rende questa specie adatta allo studio delle conseguenze genomiche subite dalle popolazioni pressate a lungo termine.
Il fagiano dalle orecchie blu ( C. auritum ) è una specie sorella di C. mantchuricum ed è originario della Cina nord-occidentale ( Zheng 2015 ; Wang et al. 2017 ). È distribuito nelle province di Qinghai, Ningxia, Gansu e Sichuan. Il C. auritum è distribuito allopatricamente con C. mantchuricum all'interno delle barriere geografiche desertiche dell'altopiano del Loess ( Zheng 2015 ). Pertanto, un flusso genico recente tra queste specie è improbabile. Un flusso genico storico tra queste specie è stato documentato e potrebbe essere stato associato alla loro divergenza relativamente tardiva (≥0,11 milioni di anni fa) ( Wang et al. 2017 ). Le popolazioni di C. auritum sono stabili e questa specie non è di interesse conservazionistico secondo la Lista Rossa IUCN ( BirdLife International 2016b ). I dati nucleari multilocus hanno rivelato un'elevata diversità genetica in C. auritum , a differenza di C. mantchuricum ( Wang et al. 2017 ). Pertanto, C. auritum è adatto al confronto con C. mantchuricum in termini di genomica della conservazione e possiamo applicare un approccio di genomica di popolazione comparativa per studiare le conseguenze del declino della popolazione in C. mantchuricum.
In questo studio, abbiamo assemblato e annotato un genoma di riferimento de novo per C. mantchuricum. Abbiamo risequenziato i genomi di 40 individui provenienti da tre popolazioni selvatiche isolate di C. mantchuricum e di 11 individui di C. auritum . Abbiamo quindi studiato la diversità genomica, il ruolo relativo della selezione purificante e della deriva genetica, la demografia storica e le conseguenze genetiche del declino della popolazione, ovvero il livello di consanguineità e il carico genetico in C. mantchuricum . Le analisi qui presentate hanno chiarito l'eredità genomica di una specie di uccello minacciata e potrebbero facilitarne la gestione della conservazione..."
Evidenza di carico genetico in C. mantchuricum
“..C'erano relativamente più mutazioni missenso in tutte e tre le popolazioni di C. mantchuricum che in C. auritum ( fig. 5 a ) . Questo risultato era coerente con la bassa diversità genetica ( fig. 2 ), la forte deriva genetica ( fig. 3 f ) e l'estesa consanguineità ( fig. 4 ) osservate in C. mantchuricum . Brown-E aveva relativamente più varianti con perdita di funzione (LOF) rispetto a Blue ( fig. 5 a ). Tuttavia, il numero di varianti LOF in Brown-C e Brown-W non differiva significativamente da quelle in Blue ( fig. 5 a ). Sebbene C. mantchuricum abbia accumulato più mutazioni missenso rispetto a C. auritum ( fig. 5 a ), il test non ha isolato le varianti deleterie e neutre. Pertanto, abbiamo calcolato il rapporto tra mutazioni missenso e mutazioni sinonime nei siti omozigoti ed eterozigoti. Si sono riscontrati tassi di mutazioni missenso significativamente più elevati nei siti eterozigoti rispetto a quelli omozigoti per tutte e tre le popolazioni di C. mantchuricum ( fig. 5 b ). Tuttavia, le mutazioni missenso si sono verificate con la stessa frequenza nei siti eterozigoti e omozigoti per C. auritum . Poiché i geni recessivi letali hanno maggiori probabilità di essere eterozigoti negli organismi viventi, questo risultato suggerisce che tutte e tre le popolazioni di C. mantchuricum hanno accumulato mutazioni recessive deleterie, mentre C. auritum no.
Abbiamo testato se la consanguineità comune a tutte e tre le popolazioni di C. mantchuricum causasse l'accumulo di mutazioni recessive deleterie. Per tutte e tre le popolazioni di C. mantchuricum , le regioni ROH presentavano meno alleli missense omozigoti rispetto alle altre regioni ( fig. 5 c ). Pertanto, la consanguineità ha effettivamente contribuito all'accumulo di mutazioni recessive deleterie. Abbiamo quindi confrontato i valori di diversità genetica per varie regioni genomiche di C. mantchuricum e C. auritum . Le regioni genomiche funzionali (complesso maggiore di istocompatibilità, elementi non esonici conservati, introni conservati, regioni intergeniche conservate e regioni codificanti) e le regioni non funzionali (presunte regioni non funzionali, introni non conservati e regioni intergeniche non conservate) di C. mantchuricum presentavano una diversità genetica inferiore rispetto alle corrispondenti regioni di C. auritum ( fig. 5 d e fig. S8 supplementare , Materiale supplementare online) . Questo risultato era coerente con l'osservazione che C. mantchuricum presentava una minore diversità genetica ( fig. 2 ). Nel complesso, questi risultati indicano che C. mantchuricum ha un carico genetico più elevato e una minore variabilità genetica adattativa.
Abbiamo esaminato geni presumibilmente funzionali al fine di valutare l'entità del carico genetico. Abbiamo selezionato 308 geni in base alla d XY dell'intero genoma e alla variazione genetica ( π ) ( figura supplementare S9 e set di dati S4 e S5, Materiale supplementare online). Due presentavano mutazioni LOF derivate e 121 mutazioni missense derivate. Una delle varianti LOF derivate era associata alla funzione di riparazione del DNA e fissata in tutte e tre le popolazioni di C. mantchuricum . Tuttavia, nessuna variante LOF derivata è stata fissata in C. auritum ( set di dati supplementare S4, Materiale supplementare online). Delle mutazioni missense derivate sui geni presumibilmente sotto selezione positiva, 48 sono state fissate in C. mantchuricum. Nove erano deleterie in base ai loro punteggi Provean. Al contrario, solo 27 mutazioni missense derivate sono state fissate sui geni presumibilmente adattativi in C. auritum e di queste, sei erano deleterie ( set di dati supplementare S5, Materiale supplementare online). I geni presumibilmente sottoposti a selezione positiva sono stati associati principalmente alla digestione, secondo le analisi del pathway KEGG e dell'arricchimento GO. La diversità genetica media di questi geni in C. mantchuricum era inferiore a quella dei geni corrispondenti in C. auritum ( set di dati supplementari S6, materiale supplementare online). Per C. mantchuricum , il tasso relativamente elevato di fissazione di mutazioni deleterie su geni potenzialmente funzionali indicava un basso potere di selezione purificante..”
Contrazione della popolazione a lungo termine in C. mantchuricum
"..Per esplorare le dinamiche delle dimensioni della popolazione di C. mantchuricum , abbiamo dedotto la storia demografica a lungo termine delle tre popolazioni utilizzando la coalescenza markoviana sequenziale multipla (MSMC). Le dimensioni effettive della popolazione ( N e ) hanno iniziato a diminuire a circa 37,5 ka ( fig. 6 ). Le N e più recenti (circa 0,5 ka) delle tre popolazioni di C. mantchuricum erano 1.420 (Brown-W), 24.041 (Brown-C) e 730 (Brown-E). Negli ultimi circa 1,64 × 10 4 anni, le dimensioni effettive della popolazione di C. mantchuricum sono sempre state inferiori a quelle di C. auritum ( fig. 6 ) . Questa scoperta è coerente con il modello di diversità genetica ( fig. 2 ). I risultati bootstrap delle fluttuazioni di N e sono mostrati nella figura supplementare S10 , Materiale supplementare online.
Inoltre, le simulazioni coalescenti eseguite in fastsimcoal26 hanno corroborato la storia della popolazione dedotta da MSMC, secondo cui la contrazione a lungo termine si è verificata in modo inclusivo nelle tre popolazioni di C. mantchuricum , sulla base degli spettri di frequenza del sito empirico ( set di dati supplementari S7 e fig. S11 , Materiale supplementare online). Inoltre, non vi era alcun flusso genico tra popolazioni adiacenti di C. mantchuricum , come suggerito dal miglior modello in fastsimcoal26, il che è coerente con la stima del flusso genico effettuata utilizzando Migrate-N ( set di dati supplementari S8, Materiale supplementare online).."
Discussione
"..Per quanto a nostra conoscenza, questo studio è il primo a condurre un'indagine genomica di popolazione di C. mantchuricum. Questa specie minacciata è ora limitata alle foreste montane della Cina settentrionale. Qui, abbiamo utilizzato un genoma de novo e dati genomici di popolazione provenienti da tre popolazioni frammentate di C. mantchuricum e da una popolazione della sua specie sorella, C. auritum . Il presente studio ha chiarito le conseguenze genomiche del declino della popolazione a lungo termine, tra cui un livello estremamente basso di diversità genetica e prove di depressione da consanguineità. L'analisi comparativa tra una specie altamente minacciata e la sua specie sorella demograficamente stabile ha dimostrato che la deriva genetica riduce l'efficacia della selezione naturale e può portare all'accumulo di mutazioni deleterie nella specie in pericolo. Nel complesso, questo studio fornisce risorse genomiche che facilitano la gestione della conservazione di C. mantchuricum .
La popolazione di C. mantchuricum ha iniziato a contrarsi durante l'ultimo periodo glaciale, circa 12,1–34,8 ka ( fig. 6 ). La traiettoria delle dimensioni della popolazione di C. mantchuricum e della popolazione cinese Han (CH) ha mostrato tendenze opposte ( fig. 6 ). Quest'ultima ha iniziato a espandersi circa 30 ka ( Schiffels e Durbin 2014 ), mentre la prima ha iniziato a contrarsi simultaneamente. Tale tendenza della popolazione di C. mantchuricum ha anche coinciso approssimativamente con il raffreddamento a lungo termine dell'altopiano centrale del Loess cinese, 34,8–12,1 ka ( Song et al. 2007 ). Tuttavia, il cambiamento climatico non è stato probabilmente l'unica ragione del declino a lungo termine di C. mantchuricum. A differenza di altre specie di fagiano come il fagiano di monte ( Tetrao tetrix ) ( Kozma et al. 2016 ), la popolazione di C. mantchuricum non si è ripresa nemmeno dopo l'inversione della traiettoria della temperatura ( fig. 6 ). La prima registrazione di attività umana nella regione geografica del presente studio risale a 39 ka ( Shang et al. 2007 ; Barton et al. 2020 ) e corrisponde approssimativamente all'accelerazione del declino della popolazione di C. mantchuricum ( fig. 6 ). Nell'ultimo millennio, l'impatto umano sulla distribuzione di C. mantchuricum è aumentato sotto forma di eventi come la costruzione della Grande Muraglia nello Shanxi, Shaanxi, Hebei e Pechino durante le dinastie Ming e Qing. Pertanto, le recenti attività antropiche in espansione come la deforestazione, i cambiamenti nell'uso del suolo e la caccia potrebbero aver contribuito al costante declino della popolazione di C. mantchuricum . La forte struttura genetica delle popolazioni isolate di C. mantchuricum ( fig. 1 ) potrebbe essere stata la conseguenza della perdita e della frammentazione di habitat idonei tra di esse, con conseguente blocco del flusso genico. Pertanto, le tre popolazioni isolate con diverso N e possono essere trattate separatamente come unità gestionali indipendenti.
Abbiamo scoperto che C. mantchuricum ha subito consanguineità durante il declino della sua popolazione poiché erano presenti modelli LD e tratti omozigoti nel suo genoma ( fig. 4 ). LD decade rapidamente a livelli molto bassi nelle popolazioni di uccelli selvatici ( Poelstra et al. 2013 ; Kawakami et al. 2014 ) come è successo per C. auritum ( fig. 4 a ) . Al contrario, C. mantchuricum ha mostrato LD estremamente elevato ( fig. 4 a ). Il declino della popolazione e la frammentazione in C. mantchuricum potrebbero aver aumentato la frequenza di consanguineità che, a sua volta, ridurrebbe ulteriormente la dimensione effettiva della popolazione. L'ROH si è verificata ampiamente nei cromosomi di C. mantchuricum ( fig. 4 b ). Gli ROH sono il risultato di alti tassi di ereditarietà di segmenti cromosomici identici per discendenza ( Ceballos et al. 2018 ). Tutte le popolazioni di C. mantchuricum presentavano frazioni di sequenze autosomiche all'interno dei ROH diverse volte superiori rispetto a C. auritum ( fig. 4 b ). Almeno metà del genoma di C. mantchuricum era contenuta in ROH lunghi e il suo tasso di LD era estremamente elevato ( fig. 4 ). Entrambi gli attributi sono conseguenze genomiche del declino della popolazione e della consanguineità, che ridurranno la diversità genetica e aumenteranno il carico genetico nelle popolazioni selvatiche.
Le firme genomiche della riduzione della diversità genetica e dell'accumulo di alleli deleteri sono state rilevate in C. mantchuricum . Rispetto a numerose altre specie di uccelli studiate utilizzando set di dati comparabili, C. mantchuricum ha una diversità genetica estremamente bassa ( fig. 2 a ). Questa scoperta era coerente con uno studio precedente che utilizzava sequenze multiesone ( Wang et al. 2017 ). In generale, la diversità genetica è negativamente correlata con l'idoneità della popolazione ( Frankham 2004 ). Pertanto, insieme alla bassa diversità genetica nella regione genomica funzionale di C. mantchuricum ( figg. 3 c e d e 5 d ), questi risultati indicano che C. mantchuricum è a maggior rischio di estinzione. Qui, abbiamo scoperto che C. mantchuricum ha accumulato più varianti missense rispetto a C. auritum e Brown-E ha più LOF rispetto a C. auritum ( fig. 5 a ). Le varianti missenso sono deleterie poiché il rapporto delle mutazioni missenso nei siti omozigoti era significativamente inferiore a quello nei siti eterozigoti ( fig. 5 b ). Pertanto, queste firme genomiche suggeriscono che C. mantchuricum stia probabilmente soffrendo di depressione da consanguineità. I modelli scoperti sono documentati in alcune specie in via di estinzione con un livello estremamente basso di diversità genetica, ad esempio la volpe delle Isole del Canale ( Urocyon littoralis ) (Robinson et al. 2016) e i cercopitechi dal naso camuso ( Rhinopithecus spp.) ( Zhou et al. 2016 ). Sorge ovvia la domanda, in popolazioni piccole e altamente consanguinee: la selezione purificante è sufficiente per eliminare le mutazioni deleterie?
La consanguineità espone mutazioni recessive letali e mutazioni parzialmente recessive dannose alla selezione purificante. In questo modo, riduce la frequenza di mutazioni dannose e il livello di depressione da consanguineità nelle popolazioni (purga genetica) ( Lande e Schemske 1985 ; García-Dorado 2015 ). Tuttavia, questa tendenza potrebbe non applicarsi a popolazioni estremamente piccole. Secondo la teoria quasi neutrale, gli alleli sono invisibili alla selezione purificante quando l'effetto selettivo è inferiore a 1/2 N e ( Ohta 1972 ; Akashi et al. 2012 ). Le popolazioni possono diminuire fino a quando la selezione purificante non elimina più le mutazioni deleterie. Studi precedenti hanno dimostrato che l'efficienza della selezione purificante varia tra le popolazioni selvatiche. Lo stambecco alpino ( Capra ibex ) ha sperimentato numerosi colli di bottiglia ma poche mutazioni altamente deleterie, il che suggerisce che la selezione ha eliminato le mutazioni deleterie ( Grossen et al. 2020 ). Al contrario, in alcuni casi si è verificata una grave depressione da consanguineità e un allentamento della selezione purificatrice per deriva genetica. Ad esempio, il carico di alleli deleteri in due sottospecie di gorilla di montagna ( Gorilla spp.) è aumentato dopo un prolungato declino della popolazione ( Xue et al. 2015 ). I risultati del presente studio sono coerenti con il caso del gorilla, indicativi di una forte deriva genetica in C. mantchuricum . Queste conseguenze genetiche dannose possono essere rafforzate se le popolazioni di C. mantchuricum continuano a diminuire. Anche se le popolazioni si riespandessero, la firma dell'erosione genetica sarebbe comunque persistente ( Kirkpatrick e Jarne 2000 ). A questo proposito, strategie di conservazione praticabili devono mirare a ripristinare le dimensioni effettive della popolazione di C. mantchuricum ( Peart et al. 2020 ).
Crossoptilon mantchuricum è attualmente elencato come "vulnerabile" dalla Lista Rossa IUCN ( BirdLife International 2016c ) ed è un animale protetto a livello nazionale di prima classe in Cina ( Zheng 2015 ). Pertanto, la caccia è severamente vietata per le popolazioni selvatiche. Per consentirne la persistenza a lungo termine, vengono proposte alcune raccomandazioni basate sui nostri risultati. Poiché dimostriamo che la selezione purificante è meno efficiente in tutte e tre le popolazioni isolate di C. mantchuricum , qualsiasi misura che favorisca l'aumento della popolazione selvatica e il ripristino di habitat idonei è molto importante, poiché la selezione funzionerebbe più efficacemente su popolazioni più grandi. In secondo luogo, poiché abbiamo trovato prove del tasso relativamente alto di fissazione per mutazioni deleterie in C. mantchuricum , il monitoraggio della salute genetica della popolazione potrebbe essere un mezzo necessario per ridurre gli effetti negativi causati dall'erosione genetica e da eventi catastrofici naturali, come malattie e incendi boschivi. Questo è probabilmente più critico per le popolazioni occidentali e orientali poiché hanno un areale geografico più piccolo e N e , confrontando la popolazione centrale nello Shanxi. Pertanto, qualora si pianificassero programmi di salvataggio genetico, come la traslocazione e la reintroduzione della popolazione, la popolazione centrale dello Shanxi potrebbe essere una popolazione di origine. Le differenze nei profili genetici tra queste tre popolazioni indicano anche la necessità di programmi di gestione della conservazione più sofisticati.."
Da: ZOOLOGICA SCIENTIFIC CONTRIBUTIONS of the NEW YORK ZOOLOGICAL SOCIETY VOLUME 30 - 1945 - Numbers 1 -18
Note riguardanti i fagiani orecchiuti (Crossoptilon) con descrizione di una nuova sottospecie Di Jean Delacour:
Mappa della distribuzione allegata alle note di Delacour esposte per esteso in questa disamina

"..Nel corso di studi approfonditi necessari alla preparazione di una nuova opera sui fagiani che il Dr. William Beebe e io speriamo di pubblicare dopo la fine della guerra, ho recentemente esaminato i fagiani orecchiuti (Crossoptilon).
Sebbene ci sia ancora molto da imparare sulla distribuzione e variabilità di questi uccelli, ora possediamo informazioni utili che non erano disponibili ventisette anni fa, quando fu pubblicata la "Monografia dei fagiani" del Dr. Beebe, e possiamo comprendere meglio l'intero gruppo. Apparentemente costituito da tre specie distinte 1*), due delle quali strettamente imparentate e altamente specializzate, mentre la terza è diversa e più primitiva.
Il fagiano orecchiuto blu (Crossoptilon auritum) mostra il più alto grado di evoluzione nella coda, normalmente composta da ventiquattro timoniere, delle quali le due paia centrali presentano membrane molto lunghe, disintegrate e ampiamente separate quasi fino all'apice. Si trova nel Gansu, zone limitrofe al Kokonor e nel Sichuan nord-occidentale. Finora è stata indicata come sua località tipo "Cina interna", che ora propongo di limitare a Si-gu, Gansu, dove Berezowski ha raccolto esemplari.
Il fagiano orecchiuto bruno (Crossoptilon mantchuricum) vive nello Shanxi settentrionale e nel Chihli 2*) nord-occidentale; è un po' meno specializzato, avendo solo 22 timoniere, la cui coppia centrale è anch'essa molto disintegrata, ma presenta una piccola macchia solida sulla punta. Limito la sua località tipo alle montagne oltre San-yu, Chihli (da A. David), in sostituzione di "vicinanze di Pechino".
Non vi è alcuna variazione geografica in queste due specie, e hanno areali completamente isolati su alte montagne. In entrambe i ciuffi auricolari sono costituiti da penne lunghe e disintegrate, e sporgono vistosamente sopra la nuca. La coda è molto compressa e solitamente tenuta verso l'alto, le timoniere centrali sono molto più in alto delle altre. Possono essere considerate come formanti una superspecie, essendo identiche per forma, voce e comportamento.
La terza specie, Crossoptilon crossoptilon, è più grande; i ciuffi auricolari sono corti e sporgono appena sopra la testa, a volte non la superano neppure. Coda composta da 20 timoniere, eccezionalmente 22, dalla struttura pressoché normale, con la metà terminale priva di lunghe membrane disintegrate. Si presenta anche molto più piatta, meno compressa e portata verso il basso. Rappresenta certamente un tipo meno specializzato e più primitivo.
L'aspetto di questa forma è sorprendentemente diverso da quello delle altre due precedenti specie, così come la sua voce e le sue azioni.
La distribuzione della specie comprese in questo genere è ancora incompleta. A differenza dei Crossoptilon mantchuricum e auritum, la specie in questione mostra una notevole variazione di colore in tutto il suo areale, che comprende le alte montagne del Sichuan centrale e sud-occidentale, dello Yunnan nord-occidentale, del Tibet orientale e del Kokonor meridionale. Nel sud-ovest, secondo Ludlow (Ibis, 1944, pp. 377-379), raggiunge Long. 91° 33' E nella valle di Tsangpo, e non si trova a sud dell'asse principale dell'Himalaya.
La specie varia di colore dal bianco quasi puro, con cappuccio e punta della coda neri, a un blu-grigio scuro.
La forma occidentale del Sichuan (Crossoptilon c. crossoptilon) è bianca con ali grigio-brunastre scure e sfumatura grigia più o meno intensa sul mantello. La coda è prevalentemente nera, con qualche soffusione grigio-brunastra sulla metà basale, riflessi verdi e viola vicino all'apice. La località tipo del Crossoptilon c. crossoptilon (Hodgson, Journal of the As. Soc. Bengal, 7, 1838, p. 864) è rimasta vaga, poiché con la descrizione non è stata specificata alcuna località. Propongo di limitarlo a Ta-tien-lu 3*) nel Sichuan occidentale, dove sono stati raccolti molti esemplari, tutti concordanti con il tipo.
La forma del Tibet orientale (Crossoptilon c. drouyni) mostra una livrea quasi completamente bianca, ed una quantità variabile, ma sempre considerevole, di bianco o grigio nella coda e pochissimo grigio sulle ali, limitato generalmente ai vessilli interni delle primarie. Si incontra e intergrada con una forma grigio chiaro (probabilmente Crossoptilon c. dolani?) nella regione dell'Alto Salween. Nel suo libro "Salween", R. Kaulback riferisce di aver trovato nel 1936 "grandi fagiani, di un bianco puro a parte gli anelli rosa intorno agli occhi e una macchia grigia sulla sommità della testa" vicino a Idashi, a nord di Zimda 4*) (vicino a 95° 30' E. Long., 31° 20' N. Lat.) e più a ovest e a nord, tra Kyierothang e Deje 5*) (94° 40' E. 31° 40' N.)" entrambe le specie completamente bianche e grigio chiaro con punte nere su ali e code, in stormi misti con tra di loro meticci di ogni tonalità". Un po' più a sud, vicino a Sating 6*) (94° 40', 31° 20'), in una rada foresta di ginepri, incontrò una cinquantina di fagiani dalle orecchie bianche e fagiani rossi (Chrysolophus pictus) che pascolavano insieme lungo il sentiero, e più a ovest, a nord del Pengar Gompa 7*) (94° 10', 31° 20') vide "altri trenta grandi fagiani, per lo più della varietà blu, con alcuni semi-nidificanti".
Non essendo stati raccolti esemplari, è impossibile sapere se gli uccelli "grigio chiaro" avvistati da Kaulback siano dolani o qualche altra forma ancora sconosciuta. Sembra improbabile che si tratti di Crossoptilon harmani, in quanto veramente molto scuro.
C'è stata molta confusione riguardo al nome da applicare alla specie più bianca, poiché Crossoptilon drouyni venne descritto da Verreaux (Nouv. Archives du Museum, 4, Parigi, 1868) da un esemplare inviato da M. Dabry, console francese a Hankow e registrato come proveniente dal "Tibet, nella parte chiamata Mu-Pin". Quest'ultima denominazione deve essere un errore, poiché il paese comunemente conosciuto come Muping 8*) è quello intorno a Ta-tien-lu dove il Crossoptilon c. crossoptilon è comunemente presente, e le serie ivi raccolte mostrano pochissime variazioni. Pertanto, la località tipo di Crossoptilon drouyni dovrebbe essere fissata e propongo di limitarla al Passo Sok, in Tibet 9*) nei pressi del quale diversi esemplari furono raccolti (incluso il tipo di leucurum) da Thorrold e Bower tra Chamdo e il Passo Sok, e dal Principe Henri d'Orléans, a ovest del Passo.
Gli individui di questa regione variano, alcuni presentano la parte basale della coda bianca e i fusti delle primarie come nel tipo maschile di Crossoptilon leucurum di Seebohm; la maggior parte di essi, tuttavia, è identica al tipo di Crossoptilon drouyni e concorda con le tavole dei Nouvelles Archives du Museum e della monografia di Elliot. Mostrano rachidi scuri sulle primarie, con vessilli interni più o meno screziati di grigio-brunastro; a volte si evince una leggerissima sfumatura grigia sul mantello e groppone, mentre a parte basale delle timoniere è grigia, con più o meno bianco sulle coppie esterne. Il l'epiteto "drouyni" di Verreaux (1868) è anteriore a quello di leucurum di Seebohm (1892) e deve essere usato per questa sottospecie.
Nel 1935, una forma grigio chiaro molto distinta, Crossoptilon c. dolani, fu scoperta dalla spedizione Dolan nel sud di Kokonor, a Jyekundo (96° 45', 33°) attuale prefettura di Yushu nel sud-est del Qinghai, e ad una certa distanza a nord sul fiume Yangtze. È stata descritta e studiata da R. M. de Schauensee ed E. Schafer (Proc. Acad. Nat. Sci. Philadelphia, 89, 1938, p. 339— Ibid, 90, 1939, p. 190). Si tratta di una variante molto chiara di Crossoptilon harmani, con le piume delle parti superiori molto ruvide al tatto, anziché lisce e piuttosto setose come nelle altre sottospecie 10*).
In generale, Crossoptilon harmani mostra una somiglianza superficiale nel colore con Crossoptilon auritum, ma si tratta di pura coincidenza, poiché le due forme differiscono nettamente sia nella forma che nelle proporzioni della coda e ciuffi auricolari, nonché ampiamente separate geograficamente. Anche nel colore, harmani differisce da auritum per il ventre bianco, fascia bianca più ampia sulla nuca e striscia bianca lungo il collo anteriore, così come per il collo e la parte superiore del dorso più scuri e marroni, la parte inferiore del dorso e groppone molto più chiari e l'assenza di bianco nella coda. È identico a Crossoptilon c. crossoptilon nella struttura della coda, dei ciuffi auricolari e nella corporatura generale. Abita la parte sud-occidentale dell'areale della specie, sui pendii settentrionali dell'Himalaya da 91° 33' a ovest nella valle di Tsangpo fino a Pome, Gyale e Pankar a ovest di Gyamda Dzong (93° 30' E, 30° 15' N).
William Beebe ha osservato e registrato fagiani dalle orecchie nello Yunnan nord-orientale e da allora sono state raccolte buone serie sulla catena del Lichiang da Forrest e da Rock. Queste serie sono abbastanza uniformi, indipendentemente dal fatto che gli uccelli siano stati catturati in primavera o in autunno; L'usura colpisce solo le parti bianche del piumaggio, che diventano giallo ocraceo a causa delle macchie molto tempo dopo la muta. Gli esemplari di Lichiang mostrano le ali più chiare rispetto al Crossoptilon c. crossoptilon, ma non bianche come nel Crossoptilon c. drouyni. Propongo per loro il nome di:
Crossoptilon crossoptilon lichiangense ( Delacour, J.T. .....1945 )
subsp. nov.
Differisce da Crossoptilon c. crossoptilon, al quale assomiglia di più, per le ali di un grigio più chiaro; vessilli esterni delle primarie grigio chiaro; secondarie e le terziarie grigio chiaro nella loro parte esposta, così come le copritrici alari. Le quattro paia esterne di timoniere presentano nella maggior parte dei casi un distinto bordo esterno grigio-biancastro, solitamente assente in Crossoptilon c. crossoptilon, e più grigio sulla metà basale di tutte le timoniere; sfumatura grigia del mantello mediamente più debole. I maschi adulti sono più chiari delle femmine e degli esemplari giovani, come in tutte le altre forme. Iride giallo oro pallido; cera grigia; becco rosa carnoso, giallastro verso la base; zampe e piedi scarlatti, artigli marrone-corno; pelle intorno agli occhi di un cremisi intenso e intenso.
Tipo 5, n. 543127, American Museum of Natural History, New York, coll., di C. Forrest, fianco orientale della catena montuosa del Lichiang, Yunnan nord-occidentale, altitudine 14.000 piedi, 10 ottobre 1922. Ali 327; coda 520; tarso 95; culmen (dalla narice) 36 mm. Otto esemplari esaminati; uno presenta 21 timoniere.
Senza dubbio, ci sono ancora molte varianti intermedie tra le diverse razze da registrare, e in molte aree esistono popolazioni instabili. Tra le forme conosciute ed esaminate, crossoptilon, lichiangense, dolani e harmani 11*) mostrano poche variazioni individuali, per contro, drouyni (=leucurum) appare piuttosto instabile in riguardo alla quantità di bianco sulle timoniere e sfumatura scura su primarie e loro peduncoli.
Ghigi ha condotto un'interessante indagine sui caratteri genetici dei fagiani orecchiuti bruni studiando e sperimentando ibridi (L’Oiseau, 1934, pp. 10-23). Sembra che da un lato le lunghe "orecchie" di auritum e mantchuricum siano dominanti, mentre dall'altro lo è anche la coda piatta, quasi normale, di crossoptilon.
Negli ibridi tra auritum e mantchuricum, il colore blu e le macchie bianche chiare sulla coda del primo sono chiaramente dominanti, mentre le timoniere centrali presentano una struttura intermedia.
I fagiani orecchiuti bruni (Crossoptilon mantchuricum) furono introdotti per la prima volta in Francia nel 1864 e allevati in gran numero, dapprima da Melle de Bellonet nel 1866, e nel 1868 e 1869 (^Bull. Soc. Accl., Parigi, 1870, p. 170) producendone diverse centinaia. Da allora, si sono ben stabilizzati in Europa e successivamente in America.
I primi fagiani orecchiuti blu (Crossoptilon auritum) arrivati vivi in Europa dalla Cina furono inviati alla mia (Delacour) collezione di Cleres nel 1929 da A. Hampe, incaricato a questo scopo. Negli anni successivi alcuni esemplari furono introdotti anche negli Stati Uniti e presto propagati. Entrambe le specie sono ora comuni in cattività, una fortuna, dato che questi grandi fagiani sono minacciati di estinzione precoce nella loro casa nella Cina settentrionale, a causa della persecuzione e della distruzione delle foreste e della macchia in cui vivono.
Alcuni fagiani dalle orecchie bianche furono inviati da Ta-tien-lu a Londra nel 1891. Una coppia depose le uova allo zoo di Berlino, ma la specie non fu mai riprodotta; solo un ibrido con il fagiano marrone fu allevato ad Anversa, e ora figura nella collezione di Tring. Venne studiato da Ghigi.
Nel 1936, diverse coppie della specie del Sichuan (Crossoptilon c. crossoptilon) furono ospitate in California dal signor W. Leland Smith, che riuscì ad allevarli con successo nel 1938 e negli successivi. Oggi negli Stati Uniti è presente un discreto numero di questi splendidi uccelli.
Il Tenente Colonnello F. M. Bailey portò in Inghilterra una coppia di Crossoptilon c. harmani qualche anno fa, ma gli esemplari morirono senza essersi riprodotti.."
1*) In passato il Crossoptilon harmani veniva, solitamente, ritenuto un ibrido, ed a seconda dell'autore, tra uno o l'altro congenere. Tuttavia studi di qualche anno fa ne suggerirono la distinzione specifica. Per contro, lavori ancora più recenti, come quello del 2015 incluso in questa disamina ne mettono, nuovamente, in discussione lo status.
2*) Il nome Zhili, alternativamente romanizzato come Chihli, indicava una regione amministrativa nel nord della Cina a partire dal XIV secolo, durante la dinastia Ming e Qing fino al 1911, quando venne convertita in provincia e ribattezzata Hebei nel 1928.
3*) Ta-tien lu, indipendentemente della trascrizione o lingua nel quale viene tradotto, corrisponde all'attuale Kangding, prefettura autonoma tibetana di Garzê, provincia del Sichuan, Cina. Storicamente, il centro urbano era noto in cinese come Dajianlu, anche Tachienlu o Tatsienlu dalla traslitterazione cinese del tibetano Dartsedo o Darzêdo .
4-5-6-7*) Località menzionate nello studio sopraesposto: N° 4 circoscritte in blu - N° 5 in rosso - N° 6 in verde - N° 7 in giallo/ocra

8 *) Muping Town è il capoluogo della contea di Baoxing nella provincia del Sichuan, Cina
9 *) Come confermato da successivi studi, uno dei quali pubblicato anche in questa disamina.
10*) Tipo di piumaggio, crespo, riscontrabile anche negli altri conspecifici. Vedere immagini nel proseguo della panoramica.
11*) Non so quanti esemplari possa aver studiato Delacour, forse non molti, sicuramente non ha avuto la possibilità, e probabilmente neppure l'ha cercata, di esaminare alcune centinaia di immagini provenienti da varie aree della gamma di queste forme. Quindi asserire che crossoptilon, lichiangense e dolani mostrano poca variazione è sostanzialmente inesatto in quanto, soprattutto dolani, perlomeno la forma attuale, mostra una notevole variazione che nella parte dedicata a questa forma, ho riassunto in...,"Cento sfumature di grigio".
Commento personale.- Trovo quantomeno azzardato, il riconoscimento specifico del Crossoptilon lichiangense attraverso i caratteri subspecifici menzionati nella descrizione. Inoltre, avendo una distribuzione (vedere mappa di Delacour) che si estende tra quelle dei congenere Crossoptilon c. drouyni e crossoptilon, si è indotti a presumere che possa essere un ibrido prodotto dal loro incrocio.
Descrizioni originali, quanto possibile, ed un poco di storia relativa alle forme comprese in questo genere:
Crossoptilon auritum ( Fagiano orecchiuto blu ) ........Pallas P.S. ....1811
Nota: non è stato ancora possibile trovare la descrizione originale del "Phasianus auritus", poi Crossoptilon auritum, di Pallas del 1811. Del resto, perlomeno secondo quanto affermato da Beebe (vedere in: un poco di storia), servirebbe solo come curiosità cronologica/storica in quanto essendo piuttosto vaga aveva indotto a non poche interpretazioni errate.
Quanto appena menzionato è ancor meglio spiegato nella successiva e breve descrizione del Crosspotilon cerulescens che, pur trattandosi senza dubbio di autitum, a causa della scarsità di dettagli forniti da Pallas, venne inizialmente scambiato per una nuova specie.
Da: THE ANNALS AND MAGAZINE OF NATURAL HISTORY, INCLUDING ZOOLOGY, BOTANY, and GEOLOGY.
VOL. V - FOURTH SERIES - 1870.
Carattere di una nuova specie di Crossoptilon. Dell'Abbé Armand Davin.
M. Milne-Edwards ha comunicato all'Accademia delle Scienze la seguente diagnosi di un Crossoptilon, estratta da una lettera di M. A. David, datata She-Tchuan, 18 dicembre 1869. La specie è denominata Crossoptilon cerulescens:
"Stesse dimensioni e forma del Crossoptilon auritum; zampe rosse; becco rosso chiaro, macchiato di marrone verso l'apice; iride rosso-nocciola; testa simile a quella della specie pechinese (Crossoptilon mantchuricum), con copritrici auricolari un poco più sviluppate; colore generale del piumaggio uniforme e molto fine, color ardesia bluastro scuro, eccetto per le estremità delle grandi penne della coda che sono nere e lucenti, con riflessi verdi e viola; le tre o quattro piccole timoniere laterali si mostrano bianche nella loro porzione basale o interamente, a seconda dell'età; anche le primarie delle ali sono color oliva; penne nere vellutate della sommità della testa separate da quelle color ardesia del collo "da una piccola striscia bianca."— Comptes Rendus, 7 marzo 1870, tomo Ixx. p, 538.
Un'altra conferma di quanto fosse difficile l'identificane specifica attraverso descrizioni non dettagliate, la offre anche Swinhoe nel 1862 che nello scambiare inizialmente, perlomeno apparentemente, una nuova forma per auritum, espone una sorta di proto descrizione, anzi, la descrizione originale di quello che dopo circa un anno verrà definitivamente riconosciuto come Crossoptilon mantchuricum.
Attenzione ! I successivi appunti descrittivi, nonostante la supposizione che costituisce il titolo dell'articolo possa fuorviare, sono da intendere come le prime note descrittive riguardanti il Crossoptilon mantchuricum.
Da: PROCEEDINGS OF THE SCIENTIFIC MEETINGS OF THE ZOOLOGICAL SOCIETY OF LONDON FOR THE YEAR 1862
In riguardo ad un esemplare che si suppone sia la femmina di Crossoptilon auritum, Pallas, proveniente dalla Cina settentrionale. Di Robert Swinhoe, F.Z.S., H.M. Console a Formosa.
Il mio amico Dr. Lamprey, del 97° Reggimento, di stanza a Tientsin (o Tianjin) mi ha inviato, mentre ero ad Amoy, l'esemplare che ora espongo, in pelle, con la dichiarazione di averne visto diversi della stessa specie, tutti simili nel piumaggio. Ho portato il mio al British Museum e, gentilmente assistito dal signor G. R. Gray, è stato confrontato con l'esemplare descritto da Hodgson proveniente dall'Himalaya. Che si tratti di specie strettamente affini è innegabile; ma sulla loro diversità ci sono pochi dubbi. Il nostro è molto più grande, con tarsi più lunghi, becco più alto alla base, gola bianca e copritrici auricolari più lunghe, piumaggio marrone scuro e coda di forma diversa. Sia dallo sviluppo della pelle guanciale che da quella del becco, così come per l'aspetto del piumaggio, si deduce che si tratti di un individuo adulto, mentre dalla forma della coda si potrebbe dedurre che si tratti di un maschio. Tuttavia, esaminando le zampe, non si evincono speroni, solo una callosità dura, come quella che contraddistingue le femmine dei Fasianidi. Sono quindi piuttosto propenso a concordare con il signor G. R. Gray nel considerarlo una femmina, ma la forma maschile della coda rappresenta una difficoltà. Confrontandolo con il Crossoptilon conservato nel Museo, è tuttavia facile notare che la coda del "pechinese" è proporzionalmente più piccola e, a giudicare dall'analogia, è piuttosto improbabile che il maschio adulto indossi una livrea così sudicia. Ora, anche supponendo che sia una femmina, non è certamente quella del Crossoptilon tibetano. Si rende quindi necessario confrontarlo con la descrizione del C. auritum esposta da Pallas nella sua "Zoografia Rosso-Asiatica", vol. II, p. 86. Pallas non fa menzione di proporzioni e misure e, inoltre, afferma che l'unica pelle ricevuta dalla Cina mancava delle zampe; ma la forma della coda, con le sue diciotto penne laterali e quattro centrali ricurve, corrisponde perfettamente, così come la gola bianca e le piume auricolari, queste ultime lunghe 3, 80 cm. Ma nel colore generale, e per molti aspetti, differisce. Il soggetto esaminato da Pallas presentava le piume nere sulla corona in nero bluastro; gola e area auricolare bianche; collo, tutto il corpo fino al groppone, insieme alle basi delle ali, in uniforme bluastro plumbeo; penne interne dello stesso colore del dorso; primarie marroni, la seconda, la terza e la quarta marginalizzate esternamente di bianco; coda con le quattro timoniere centrali ricurve e vellutate, di un nero bluastro. Le quattro penne vettoriali più vicine su ciascun lato sono più larghe e intere, curvate verso l'interno e di lunghezza quasi uguale, blu alle estremità, mentre le restanti penne laterali diminuiscono gradualmente di lunghezza, la maggior parte delle metà basali è bianca, le porzioni apicali sono nero-bluastre.
Il nostro, proveniente da Pechino, presenta le piccole piume sulla sommità di colore nero-violaceo, bordate da una fascia occipitale biancastra indistinta. Gola ed area auricolare bianche. Collo in nero profondo e lucente. Dorso, ventre e ali intere marrone cioccolato intenso; addome setoso e più chiaro. Groppone e coda bianco sporco, rachidi e calami delle penne caudali in marrone cioccolato intenso, le estremità delle quali più o meno profondamente orlate di nero-violaceo, con le quattro centrali si presentano alquanto scomposte, mentre le altre nove su ciascun lato sono quasi ugualmente graduate e curvate verso l'interno.
Ora, le obiezioni che muovo contro il considerare questo esemplare la femmina di Crossoptilon auritum derivano innanzitutto dal suo tipo di colorazione. Il maschio di questa forma mostra l'intero piumaggio bluastro plumbeo. Nel nostro soggetto, di conseguenza, se si tratta di una femmina della stessa specie, potremmo aspettarci di trovare un marrone uniforme. Per contro, il nostro è caratterizzato dall'avere il collo nero e groppone bianco. I margini bianchi delle penne potrebbero essere una differenza sessuale; però colpisce, dalla descrizione di Pallas, che il nostro abbia ali molto più piccole e di forma leggermente diversa. Anche nella coda dovremmo aspettarci una maggiore somiglianza cromatica, se non di forma. Nella colorazione della coda, C. auritum si avvicina di più al C. tibetanum, Hodgson. Le quattro penne centrali sono nero bluastro; le quattro successive su ciascun lato, di lunghezza quasi uguale, presentano apici blu; mentre l'intera coda del nostro esemplare è bianco sporco, con vertice nero violaceo, e le quattro penne adiacenti a quella centrale sono graduate più o meno nella stessa proporzione di quelle che seguono. Ritengo, quindi, dopo debita riflessione, che il nostro soggetto, oltre ad esibire valide ragioni per essere considerato una femmina, sia una specie cui maschio potrebbe essere più bello del C. tibetanum e auritum. Se sono correttamente informato, l'esemplare da noi esaminato arriva dalla Manciuria 1*) mentre quello descritto da Pallas proveniva dalla Mongolia 2*) e la pelle studiata da Hodgson dal Tibet. Per il momento, quindi, mi approprio del vantaggio del dubbio e propongo di presentare questo esemplare come la femmina di una nuova specie, che propongo di chiamare Crossoptilon mantchuricum.

Manciuria: in rosso scuro = Manciuria interna - medio = Regione settentrionale della Mongolia Interna - chiaro = aree cedute alla Russia nel 1858 e nel 1860 (a volte chiamate Manciuria esterna).
Commento personale:
1-2*) Entrambe le località necessitano di un chiarimento. Il fagiano orecchiuto bruno non si trovava e trova in Manciuria, tuttalpiù si potrebbe disquisire asserendo che ne lambisce i confini meridionali nelle vicinanze di Pechino. Quindi l'epiteto è improprio. Sicuramente il Crossoptilon auritum non viveva, e vive, nella Mongolia quand'anche, come tale, fosse intesa l'attuale provincia cinese della Inner Mongolia. Non vi era nessuna certezza riguardante l'origine della pelle studiata da Hodgson, quindi Tibet dovrebbe essere inteso nel senso più ampio del termine coinvolgendo l'intero altopiano dove, tra l'altro, albergano tutte le sottospecie di Crossoptilon crossoptilon.
Descrizione e info da: A "MONOGRAPH of the PHEASANTS" del 1918 Vol. 1 di William Beebe
Un poco di storia:
Il fagiano orecchiuto blu fu il primo del suo genere ad essere descritto ma, curiosamente, è sempre rimasto il meno conosciuto tra tutti i congenere. Nel 1811 Pallas lo descrisse attribuendogli il nome di Phasianus auritus nella sua "Zoografia Rosso-Asiatica". Il tipo è andato ormai perduto da tempo e la descrizione originale era così poco dettagliata che, quando furono scoperti esemplari della forma bruna (Crossoptilon mantchuricum), le due specie furono confuse per molti anni. Tuttavia, quando l'Abbé David inviò due pelli di orecchiuto blu (Crossoptilons auritum) al Museo di Parigi, ogni dubbio fu dissipato sulla relativa distinzione delle due forme.
Maschio adulto. - La sommità della testa è ricoperta da piume corte, nere, ricurve e vellutate; le remiganti e i lati della fronte, il mento, la gola e le copritrici auricolari molto allungate sono bianchi. Tutto il piumaggio superiore e inferiore è grigio-bluastro, con la maggior parte delle membrane disintegrate, lasse e pelose. Le prime file di vere penne di contorno sul dorso occipitale della corona vellutata sono bianche, formando una stretta fascia trasversale di questo colore che si estende da una copritrice all'altra. Remiganti secondarie marrone scuro, con una forte lucentezza viola. Remiganti primarie marrone opaco più chiaro, non lucide.
Coda tipicamente con ventiquattro timoniere, le due paia centrali grigio-bluastre con le membrane completamente disintegrate fino al fusto. Le barbe sono molto lunghe, ricurve e simili a peli. Verso l'estremità diventano più scure e fortemente lucide di un verde metallico, virando all'apice al viola, con una piccola area spatolata di solida membrana, fortemente curvata verso il basso. Le successive paia di timoniere sono più saldamente palmate, con la membrana esterna di un verde fortemente iridescente e quella interna viola-viola. Le cinque o sei paia esterne di penne della coda mostrano una quantità variabile di bianco basale; negli esemplari tipici, tre quarti dell'area basale sono di questo colore, mentre la porzione distale delle penne è di un viola metallico.
Pelle facciale nuda, scarlatta; iridi, giallastre; mandibole, corno rossastro; zampe e dita, scarlatte; speroni, più chiari; artigli, corno scuro. Lunghezza: 96,0; becco dalla narice: 3,0; ala: 30,6; coda, 56,0; tarso, 10,1; dito medio e artiglio, 8,0 mm. Speroni, corti, robusti, conici, lunghi da 0,7 a 1,2 cm.
Femmina adulta. - Assomiglia al maschio, ma con gli speroni rudimentali, ed è di dimensioni leggermente inferiori. Lunghezza del becco: narice 28; ala 290; coda 490; tarso 94; dito medio e artiglio 71 mm.
Da: FIELDIANA – ZOOLOGY A continuation of the ZOLOGICAL SERIES of FIELD MUSEUM of NATURAL HISTORY CHICAGO, U.S.A. - VOL. 53, N° 1 - 27-01-1967
By MELVIN A. TRAYLOR, Jr.
Crossoptilon auritum (Pallas)
1 Femmina 1 Maschio, Sungpan, 10 e 27 ottobre 1934.
Delacour (1951, p. 199) afferma che l'areale del Crosoptilon auritum è limitato a Kokonor e Kansu e che le segnalazioni del Sichuan nord-occidentale sono probabilmente errate. Nella sua mappa (p. 185) della distribuzione del genere, lascia un intervallo di diverse centinaia di miglia tra gli areali di Crossoptilon auritum e Crossoptilon c. crossoptilon. Sulla base degli esemplari attuali e delle segnalazioni precedenti, su cui vi sono davvero pochi motivi di dubbio, gli areali di auritum e crossoptilon sembrano incontrarsi alla latitudine di Sungpan nel Sichuan settentrionale(Cina)
Weigold (in Jacobi, 1924, p. 4) considerava auritum raro a Sungpan. Non vide uccelli vivi, ma trovò le loro piume intorno a Sungpan e acquistò un adulto che si presumeva fosse stato catturato a Matschou e diversi pulcini a Hwanglungsze. Stone (1933, p. 179) registra il Crossoptilon auritum da Karlong, circa 80 km a ovest-nord-ovest di Sungpan, e Crossoptilon c. crossoptilon da Datsang, circa 160 km a ovest-sud-ovest. Dolan (in Stone, 1933, p. 167), descrivendo l'itinerario della sua spedizione, osserva che Datsang si trova sullo spartiacque tra i fiumi Hwang-ho (Fiume giallo) e Yangtze (Fiume azzurro), e che i fagiani orecchiuti a nord dello spartiacque, a Karlong e Merge, erano auritum, mentre a sud si trovava il crossoptilon. Infine, Smith o i suoi raccoglitori catturarono entrambe le specie nelle vicinanze di Sungpan. Sembra esserci poca ragione di dubitare che gli areali di queste due specie si incontrino tra i 32° e i 33° di latitudine nord.
Litografia colorata a mano con Crossoptilon auritum realizzata da Joseph Wolf e J. Smit (1870-72) per Daniel Giraud Elliot

Crossoptilon auritum ( Fagiano orecchiuto blu ) ........Pallas ......1811
Caratteristiche subspecifiche e info: la lunghezza media si aggira sui 98 cm con un peso di circa 2 kg. La livrea appare uniformemente blu-grigia; i lati della testa cremisi; ciuffi auricolari bianchi che sporgono sulla parte superiore della nuca; penne della coda centrali lunghe e rivolte verso l'alto, decomposte e cadenti; timoniere più esterne bianche. Dimorfismo sessuale pressoché inesistente con femmina e maschio sono molto simili sia nella forma del corpo che nel colore del piumaggio. Tuttavia il maschio, oltre ad essere di norma leggermente più grande, presenta uno sperone per tarso assente nella femmina.
Endemico della Cina è distribuito nel Qinghai nord-orientale e orientale, Monti Qilian meridionali nel Gansu nord-occidentale, nei Monti Helan nel Ningxia e Sichuan settentrionale. Abita praterie rade, foreste di pini o querce nelle valli a un'altitudine di circa 3.000 metri. Fatta eccezione per la stagione riproduttiva, vive spesso in gruppi di 10-30 esemplari, ma può raggiungere anche più di 100.
Le attività di foraggiamento si svolgono principalmente al mattino e pomeriggio, con maggiore frequenza al mattino presto e sera. Si riproduce da aprile a luglio. Nidifica sotto fitti cespugli o in siti leggermente concavi. Il nido è estremamente semplice a forma di disco, generalmente costituito da foglie, rami, paglia, muschio, ecc. e imbottito di piume. La deposizione consiste in 6-12 uova di colore grigio-bluastro lucido, grigio-marrone o verde chiaro, con occasionali macchie marrone chiaro incubate per 25-28 giorni. I pulcini maturano precocemente. Parrebbe che, appena schiusa, la femmina sia leggermente più grande del maschio con un peso di circa 30-32 grammi e lunga 105-110 mm. Entro i 90 giorni di età, peso e lunghezza aumentano rapidamente. A 120 giorni di vita, il becco sia nei maschi che nelle femmine supera i 30 mm ed il tarso oltre gli 84 mm.; a 150 giorni di età, la lunghezza del corpo, quella delle ali e coda raggiungono le dimensioni adulte, dopodiché la coda della femmina smette di crescere, mentre quella del maschio aumenta leggermente. La durata della vita può raggiungere i 20 anni.
Il 5 febbraio 2021 è stato inserito nel secondo livello della "Lista nazionale delle specie selvatiche protette" della Cina. Secondo i documenti storici, in passato gli uccelli nel Ningxia prosperavano. Durante il periodo Kaiyuan della dinastia Tang, le "piume dei fagiani blu" erano uno dei tributi del Ningxia. La prima testimonianza di fagiani blu è apparsa nei "Nuovi documenti del Ningxia nel periodo Jiajing", a indicare che questo rara forma ha una storia di sopravvivenza sui Monti Helan di almeno 500 anni.
Il fagiano orecchiuto blu è stato classificato come vulnerabile nell'edizione del 1998 della " Lista rossa degli animali in via di estinzione in Cina " com'anche in quella del 2009. Nel 2016 è stato inserita nella Lista Rossa IUCN delle specie minacciate con una classificazione di rischio minimo (LC) ed è stato aggiunto all'elenco degli animali selvatici protetti di secondo livello della " National Key Protected Wildlife List " della Cina nel 2021.
Foto da: 1 www.media.gzstv.com Autore: Wu Weidong - 2 www.birdnet.cn Autore: Yan Jun - 3 www.gegdjq.com ( località indicate dai fotografi: 1 solo Gansu 04-03-2025 Cina - 2 nessuna info - 3 Contea di Tanchang o Dàngchāng, Gansu 23-08-2021 Cina )
1 - Crossoptilon auritum 2 - Crossoptilon auritum 3 - Crossoptilon auritum



Crossoptilon auritum ( Fagiano orecchiuto blu ) ........Pallas ......1811
Foto da: 1-2 www.media.gzstv.com Autore: Wu Weidong ( località indicate dai fotografi: 1-2 solo Gansu 04-03-2025 Cina )
1 - Crossoptilon auritum 2 - Crossoptilon auritum
.jpg)

Crossoptilon c. crossoptilon
Descrizione originale del (Phasianus (Crossoptilon) crossoptilon di Brian Houghton Hodgson del 1838
Da: "THE JOURNAL OF THE ASIATIC SOCIETY OF BENGAL" : EDITED BY JAMES PRINSEP, F.R.S VOL. VII.—PART II. JULY TO DECEMBER, 1838.
Asiat. Soc. Beng., 7 : 864 & pl. Holotype, Adult male (relaxed mount). Reg. no. 1859.3.4.601. Locality unknown. Collected by a Nepalese envoy who was returning from Peking, and obtained from him in Nepal by Hodgson. Hodgson Collection.
(The plate is captioned Phasianus (Crossoptilon tibetanus)
On a new species of Pheasant from Tibet. By B. H. Hodgson, Esq.
Phasianidae o Pavonidae.
Genus—new ? Crossoptilon*, nob. Type Phasianus crossoptilon, nob. Hab. Tibet.
Disegno allegato alla descrizione originale del 1838 di Hodgson.

Resta solo da notare la livrea di questa forma, che costituisce in effetti la sua caratteristica più notevole. Il piumaggio, quindi, su tutto il corpo è molto ampio, non lucido, e completamente arruffato tanto da ricordare la famiglia degli Struthious. Questa peculiarità ha suggerito il nome che ho applicato all'esemplare esaminato: un nome che, per il momento, può essere considerato specifico, ma soggetto a promozione a rango generico o sub-generico, se si dimostrasse che la forma è tipica e non semplicemente aberrante. Al momento propendo a considerarlo come da prima ipotesi assegnando al tipo un posto tra Phasianus ed Euplocomus o Nycthemerus, un tipo che, tra l'altro, ho caratterizzato 11 anni fa sull'Oriental Quarterly, con lo stile di Gallophasis, indicando come icona il Kalij del Nepal di Kirxpatricx.
"..Possiedo un solo esemplare di questa grande e appariscente forma. È un maschio adulto, ed è stato portato di recente a Cathmandu (Nepal) dall'inviato nepalese a Pechino, appena rientrato. La lunghezza, dalla punta del becco a quella della coda, è compresa tra 97 e 102 cm., dei quali il becco è lungo 3,80 cm., e la coda tra 49 e 51 cm.
L'ala chiusa misura 31,75 cm.; tarso 11,43 cm., ed il dito centrale 6,35 cm. Il becco ha la stessa lunghezza, sia preso dall'apertura che dalla fronte, ed è 0,9525 cm., più corto della testa in totale lunga 5,8 cm. Il becco molto robusto, mostra le caratteristiche generali di quello del Lophophorus, con il bordo tomiale 1*) della mandibola superiore ancora più smussato e munito di un piccolo festone a forma di dente. La sua base è nuda. Testa e gola ricoperte di piume, tuttavia le guance, dalle narici all'occipite, ne sono prive di piume, essendo contraddistinte dalla tipica pelle rossa e papillata presentata dalla tribù dei fagiani, e in tutto quel grado di sviluppo che caratterizza più particolarmente i Kaliches indiani (Leucomelanus), le specie dipinte 2*) e Amherstiane della Cina. Come il vero fagiano (Colchicus), il nostro esemplare è privo di cresta, sebbene le piume che occupano la sommità della testa siano di tipo peculiare, essendo corte, vellutate, spesse, erette, con le punte leggermente scomposte e quadrate leggermente ricurve in avanti.
Le ali non presentano particolarità. Sono corte, rigide, arcuate e arrotondate, e come di consueto la sesta penna è la più lunga. La coda, molto ampia, è notevole soprattutto per l'ampiezza delle piume. La sua lunghezza è moderata, e non presenta l'allungamento ed il restringimento eccessivo delle penne centrali, che caratterizzano i fagiani tropicali. Composta da 18 penne 3*) regolarmente e considerevolmente graduate, con forma generale ampiamente convessa, senza alcun sintomo di compressione e curvatura tipica della Gallus. Zampe e piedi sono ben adattati al movimento rapido sul terreno e hanno forma e proporzioni molto simili a quelle di Leucomelanus e T. satyra. I tarsi sono nudi e bi-squamati anteriormente e posteriormente: ma le squame posteriori sono più piccole di quelle anteriori. I lati dei tarsi sono papillo-reticolati. Lo sperone è aguzzo e curvo. Le dita laterali sono uguali; la centrale è lunga mentre la posteriore è corta e sollevata, come di consueto. Le unghie sono lunghe e presentano una curvatura minima.
Crossoptilon tibetanus - in: THE GAME BIRDS of India, Burmah & Ceylon by Hume & Marshal Vol. II del 1879. Elaborazione del disegno originale di Hodgson realizzata da Hume 4*)
Compressione obliqua e curvatura della coda costituiscono il carattere principale di quel tipo (Gallophasis o Euplocomus) e, poiché è un carattere destinato a scomparire nella pelle secca, non sono del tutto certo che il nostro soggetto attuale non lo possegga allo stato vivo. Se così fosse, questa forma sarebbe ascrivibile ai Gallophasis o Euplocomus, ma in caso contrario, un tipo vicino, affine al vero fagiano per l'assenza di cresta, e distinto tra tutti i suoi congeneri per l'ampio piumaggio frangiato trasmesso anche alle penne centrali della coda, che mostra vessilli molto ampi ed uniformi ben separati e curvati verso l'esterno, oltre ad essere ornati da una fine lucentezza.

Il colore generale è un canuto bluastro, più chiaro e tinto di giallo sulla superficie inferiore: sommità del capo nera e vellutata; grandi piume alari e caudali scure o nere, più o meno lucide di un blu cangiante, in particolare le penne della coda; zampe e guanciali di un intenso sanguigno; becco rosso ocra opaco; iride marrone.."
Nepal, settembre 1838.
1*) Dente tomiale, o la tomia mandibolare: non è un vero dente, ma una sorta di protuberanza visibile sul bordo esterno dei becchi.
2*) Non è ben chiaro cosa intenda Hodgson con "specie dipinte", considerato che in Cina, le forme definibili "dipinte" sono moltissime.
3*) In questa specie le penne della coda sono solitamente 20.
4*) Forse la maggiore conoscenza che avevano di queste forme all’epoca in cui è stata prodotta l’elaborazione, probabilmente realizzata non sul modello della pelle originale, abbia indotto, o meglio, influenzato in modo non corretto l’illustratore. Infatti, il fenotipo raffigurato sembra quello del Crossoptilon c. crossoptilon, mentre l'esemplare descritto da Hodgson doveva corrispondere alla forma successivamente decritta come Crossoptilon c. dolani.
Da: A HAND-BOOK TO THE GAME-BIRDS ZOOLOGICAL DEPARTMENT, BRITISH MUSEUM, VOL. 1. SAND-GROUSE, PARTRIDGES, PHEASANTS. LONDON : 1895 By W. R. OGILVIE-GRANT.
Crossoptilon tibetanum ( Fagiano orecchiuto di Hodgson )
Phasianus {Crossoptilon) tibetanus Hodgson, J. As. Soc. Beng. vii. p. 864, pi. 46(1838); id. Ind. Rev. iii. p. 593, pi. (1839) Crossoptilon tibetanum, Sclater, List of Phasian. p. 6, pi. 4 (1863); Elliot, Monogr. Phasian. i. pi. 14 (1872); David and Oustalet, Ois. Chine, p. 407, pi. 107 (1877); Hume and Marshall, Game Birds of India, i. p. 115, pi. (1878); Ogilvie-Grant, Cat. B. Brit. Mus. xxii. p. 293 (1893). Crossoptilofi auritum, G. R. Gray {/lec Pallas), Gen. B. iii. p. 495, pi. cxxv. (1845). Crossoptilon drouynii, Verreaux, N. Arch. Mus. Bull. iv. p. 85, pi. iii. (1868); Elliot, Monogr. Phasian. i. p. xviii. pi. 15 (1872).
Maschio adulto. — Corona ricoperta di piume nere corte, morbide e arricciate; lunghi ciuffi auricolari bianchi 1*) come in tutte le altre specie. L'intero piumaggio superiore si presenta bianco puro, sfumato di grigio sulle copritrici alari e caudali più lunghe; penne più grandi brunastre; coda con venticinque timoniere, nere, lucide di blu-verdastro scuro e viola intenso verso l'estremità A*). Lunghezza totale: 91; ala: 31,4; coda: 47,25; tarso: 9,9 cm.
Femmina adulta. — Piumaggio perfettamente simile, ma leggermente più piccola e priva di speroni.
Gamma. — Montagne della Cina occidentale e del Tibet orientale 2*) L'esemplare tipico descritto da Hodgson fu portato in Nepal da un inviato che era stato a Pechino, ma l'esatta località in cui fu trovato l'esemplare in oggetto non venne mai accertata.
Abitudini. — Questo splendido fagiano bianco vive nelle pinete ad altitudini che variano dai 3000 ai 3700 metri sul livello del mare. È estremamente socievole e si dice che se ne possano trovare quaranta o cinquanta esemplari, appollaiati insieme sui rami dei pini.
L'abate David ci informa che questo Crossoptilon bianco si incontra solo in alcune località boscose della Cina, sulle alte montagne del Sichuan occidentale, nei pressi di Moupin e Ta-tsien-lou, dove la sua esistenza è protetta dal rispetto superstizioso degli indigeni. È un uccello molto gregario, che ama vivere in compagnia di molti suoi simili, anche quando è impegnato ad allevare i piccoli, e non si allontana molto dal luogo di riproduzione. Si nutre di foglie, radici, cereali e insetti. Fortunatamente per la sua sicurezza, la carne di questo fagiano dalle orecchie è moderatamente buona da mangiare e i cacciatori preferiscono i fagiani più piccoli (Phasianus) come selvaggina, poiché sono più ampiamente diffusi e più facili da reperire.
A*) Nel tipo di Hodgson le sei paia esterne di penne della coda presentano una macchia bianca oblunga sulla membrana esterna che corre quasi parallela allo stelo, ma queste marcature non sono simmetriche sui due lati e, in tutti gli altri esemplari che abbiamo esaminato, sono completamente assenti.
1*) In questa specie i ciuffi auricolari non oltrepassano la linea superiore del cranio, tuttavia non sembra che sia gli autori, quanto gli artisti che la raffigurano, ne prendano atto. E ciò è piuttosto strano.
2*) Pur essendo del tutto presunta, casualmente potrebbe corrispondere alla realtà dato che l'esemplare descritto da Hodgson, con ogni probabilità, come anche citato da Bebee (vedere a seguito) presentava le timoniere esterne macchiate di bianco, ali con apici nerastri, e piumaggio arruffato, quindi doveva trattarsi del Crossoptilon dolani o di uno degli ibridi, chiari, che albergano nelle aree tibetane presupposte.
Crossoptilon c. crossoptilon
Da: "A MONOGRAPH of the PHEASANTS" del 1918 by Beebe, William
Sinonimi:
Phasianus (Crossoptilon) tibetanus Hodgson, J. As. Soc. Beng., VII. 1838, p. 864, pl. 46; id. Ind. Rev., III. 1839, p. 593.
Crossoptilon tibetanum Hodgson, in Gray’s Zool. Misc., 1844, p. 85; Sclater, List of Phas., 1863, p. 6, pl. 4; Gray, List Gallinae Brit. Mus., 1867, p. 31; David, N. Arch. Mus. Bull., VII. 1871, p. 11 [Moupin]; Elliot, Mon, Phas., I. 1872, pl. 14; David and Oustalet, Ois. Chine, 1877, p. 407, pl. 107 [W. Sze-chuen]; Hume, Stray Feathers, VII. 1878, p. 426; Scully, Stray Feathers, VIII. 1879, p. 343; Seebohm, Ibis, 1891, p. 378; Sclater, Proc. Zool. Soc., 1891, p. 464; Seebohm, Bull. Brit. Orn. Club, I. 1892, p. XVIII; Oustalet, Ann. Sci. Nat., (7), XII. 1892, p. 315 [part]; Ogilvie-Grant, Cat. Game-birds Brit. Mus., XXII. 1893, p. 293; Bower, Journey across Tibet, 1894, p- 294; Ogilvie-Grant, Hand-book Game-birds, I. 1895, p. 252; Sclater, List Animals London Zool.- Gardens, 1896, p. 486; Davies, Ibis, 1901, p. 409; Tegetmeier, Pheasants, 1904, p. 228; Mitchell, Proc. Zool. Soc., 1911, p- 521; Beebe, Zoologica, I. No. 15, p. 275.
Crossoptilon auritum Gray, Genera Birds, III. 1845, p. 495, pl. CXXV; id. Cat. Hodgs., ed. I. 1846, p. 124.
Crossoptilon drouynii Verreaux, N. Arch. Mus. Bull., [V.1868, p. 85, on III [Moupin]; Swinhoe, Proc. Zool. Soc.,1871, p. 399; Elliot, Mon. Phas., I. 1872, p. XVIII, pl. 15.
Crossoptilon tibetanus Gray, Eanes list Birds, II. 1870, p. 259.
Crossoptilum tibetanum Hume and Marshall, Game-birds India, I. 1878, p. 115, pl.; Seebohm, Ibis, 1891, p. 378 [W. Sze-chuen]; Blanford, Fauna Brit. India, Birds, IV. 1898, p. 88; Sharpe, Hand-list Birds, I. 1899, p. 35; Oates, Cat. Eggs Brit. Mus., I. rgo1, p. 53, pl. V, fig. 4; Dresser, Man. Palae. Birds, 1903, p. 671.
Crossoptilon leucurum Seebohm, Bull. Brit. Orn. Club, I. 1892, p. xvii; id. Ibis, 1893, p. 250; Bower, Journey across Tibet, 1894, p. 296.
Crossoptilum leucurum Dresser, Man. Palae. Birds, II. 1903, p. 671.
Crossoptilun tibetanum Wilson, A Naturalist in Western China, II. 1914, p. 122.
Crossoptilon tibetanum tibetanum Baker, Jour. Bomb. Nat. His. Soc., XXIV. 1916, p. 626.
Crossoptilon tébetanum drouyni………………………………………………........................1916, p. 629.
Storia "antica": Hodgson, nel 1838, descrisse un fagiano bianco con il nome di Phasianus tibetanus 1*). Ottenne questa forma in Nepal da un inviato indigeno appena tornato da Pechino e, poiché per molti anni non vennero procurati altri esemplari, i suoi habitat rimasero per un periodo di tempo altrettanto lungo completamente avvolti nel mistero. Alla fine si scoprì che il tipico tibetanus non mostrava macchie bianche sulle penne della coda, e quindi l'uccello di Hodgson non era assolutamente purosangue, ma presentava una certa traccia di auritum.
Maschio adulto.- La sommità della testa è ricoperta da piume corte nere, arricciate e vellutate. Copritrici auricolari bianco-argenteo, molto allungate, che formano un lungo ciuffo bianco 2*), ricurvo verso l'alto su ciascun lato della testa. L'intero piumaggio, sia della parti superiori che inferiori, è bianco puro, sfumante in grigio sulle copritrici alari più lunghe e parte superiore della coda, con la struttura delle penne estremamente lassa e pelosa; secondarie bruno-nerastre, leggermente iridescenti di porpora; primarie marrone scuro. Coda composta da venti penne, di colore bronzo-violaceo verso la base, che sfumano in blu-verdastro scuro e viola intenso verso l'estremità delle penne. Mentre le porzioni marginali delle barbe di queste penne centrali sono lunghe, ricurve e molto lasse, non si osserva la disintegrazione tipica delle timoniere centrali del mantchuricum e auritum, che sembrano essere dovute a brusche mutazioni di lunghezza delle copritrici superiori della coda.
Parti nude ai lati della testa; zampe e piedi scarlatti; becco corno rossastro; iride giallo-arancio. Lunghezza: 92 cm; becco dalla narice: 3,8 cm; ala: 33 cm; coda: 57,5 cm; tarso: 10 cm; dito medio e artiglio: 8,1 cm. Gli speroni sono robusti, corti e conici.
Tra i fagiani orecchiuti bianchi adulti puri esiste una grande variazione nella tonalità esatta o nel grado di bianco. Impiegano senza dubbio diversi anni per eliminare completamente il pigmento marrone che predomina nel piumaggio immaturo. Della livrea, il mantello sembra mantenere la sfumatura grigia più a lungo delle altre parti, ma le primarie sono l'indicatore migliore. Negli esempi estremi di bianco (uccelli provenienti dall'estremo ovest, lontani dai luoghi di auritum), i vessilli esterni delle primarie sono bianco puro mentre quelle interne grigio chiaro. Questi individui mostrano il piumaggio bianco rosato, per contro, gli esemplari meno estremi presentano una sfumatura più tendente al crema.
Negli esemplari rosati del Tibet occidentale, le penne della coda sono tutte grigie alla base, sfumando in un viola iridescente sulle esterne. Procedendo verso l'interno, appare una sfumatura verde bronzo, e sulle timoniere centrali questo colore occupa tutta la porzione mediana dei vessilli tra la base grigia e il terzo terminale viola.
Femmina adulta.- Simile nel piumaggio al maschio, ma di dimensioni leggermente inferiori. Becco dalla narice, 2,7; ala, 29,8; coda, 40; tarso, 9,7; dito medio e artiglio, 7,6 cm.
Litografia colorata a mano con Crossoptilon tibetanum (Crossoptilon c.. crossoptilon) realizzata da Joseph Wolf e J. Smit (1870-72) per Daniel Giraud Elliot con ciuffi auricolari raffigurati non correttamente.

Crossoptilon c. crossoptilon - www.douyin.com Autore: Shǐzhōng xiāngxìn (località indicata: Jiagenba, Kangding, Ganzi, Sichuan 03-10-2023 Cina )

1*) Da quanto è in mia conoscenza, Hodgson descrisse questa forma attribuendole il nome di Phasianus crossoptilon, stabilendo anche un nuovo genere, appunto il Crossoptilon. In nessuna parte della sua descrizione, sopra esposta, eccetto l'illustrazione, si legge come nome alternativo l'epiteto "tibetanus", ricorrente invece nei lavori dei suoi colleghi che, probabilmente, ritennero più consono il binominale Crossoptilon tibetanus al Phasianus crossoptilon.
2*) In questa specie i ciuffi auricolari non oltrepassano la linea superiore del cranio. Quindi Beebe, così Joseph Wolf e J. Smit (1870-72) per Daniel Giraud Elliot, sbagliano a raffigurarla con coperture auricolari esageratamente pronunciate.
Crossoptilon c. crossoptilon ( Fagiano orecchiuto bianco ) .... Hodgson.....1838
Da: www.datazone.birdlife.org 2024
Il Crossoptilon ccrossoptilon è presente in Cina, nelle regioni del Qinghai, Sichuan, Yunnan e Tibet. Occasionalmente si è ipotizzata la sua presenza nell'India nord-orientale (ad esempio, Rasmussen e Anderton 2012), tuttavia la sua distribuzione conosciuta non si avvicina particolarmente al confine indiano e neppure ci sono conferme della sua presenza. Analogamente, mancano prove dell'esistenza di questi fagiani nel Myanmar nonostante l'areale della specie si avvicini particolarmente.
Abita foreste di conifere e miste vicino al limite degli alberi, oltre a macchie di betulla subalpina e rododendro, a 3.000-4.300 m (occasionalmente fino a 2.800 m in inverno). La sua distribuzione sembra essere determinata principalmente dall'acqua, dai siti di foraggiamento e dalla predazione (Fei Jia et al. 2005). Può essere comune intorno ai monasteri buddisti dove riceve protezione culturale, ma generalmente si trova a densità molto basse in piccoli gruppi (A. Pack-Blumenau in litt . 2006). La specie è monogama durante la stagione riproduttiva (Wu Yi e Peng Jitai 1996), anche se occasionalmente può riunirsi in gruppi fino a 30 individui nel corso dell'inverno. Si nutre beccando il terreno e scavando per trovare i bulbi 1*).
L'area di habitat idoneo è vasta all'interno della sua gamma, nella quale la specie si riscontra spesso come comune/regolare (ad esempio Madge e McGowan 2002, Brooks et al . 2019, Li et al . 2022, eBird 2024). Sebbene la caccia sia considerata una minaccia locale, essendo gran parte della distribuzione di questa specie remota potrebbe plausibilmente costituire una minaccia in grado di sopprimere globalmente i numeri a una densità sufficientemente bassa da sospettare che la dimensione della popolazione possa essere limitata. Di conseguenza, sebbene la densità della popolazione non sia formalmente stimata in questo caso, si ritiene altamente probabile che si tratti di decine, se non centinaia, di migliaia di esemplari.
Le due principali minacce per questa specie sono la perdita di habitat e la caccia. Nessuna delle due sembra rappresentare una minaccia significativa per lo status globale della specie e probabilmente non ne sta causando il declino, dato che gran parte dell'areale di questa specie è abbastanza lontana dallo sfruttamento umano.
Questa specie occupa un areale molto vasto, pertanto non si ritiene che si avvicini alle soglie di Vulnerabilità secondo il criterio di estensione dell'areale (estensione di presenza <20.000 km² combinata con un areale in calo o fluttuante, estensione/qualità dell'habitat o dimensione della popolazione e un numero limitato di località o una forte frammentazione). Si sospetta che il trend sia stabile, pertanto le soglie relative al criterio di tendenza della popolazione (declino >30% in dieci anni o tre generazioni) non sono raggiunte. La dimensione della popolazione non è stata quantificata, ma non si ritiene che si avvicini alle soglie di Vulnerabilità secondo il criterio di estensione della popolazione (<10.000 individui maturi con un declino continuo stimato >10% in dieci anni o tre generazioni, o con una struttura di popolazione specifica). Per questo motivo, la specie è stata classificata come a Minore Preoccupazione.
Commento personale: 1*) Studi sul campo condotti in Cina, così come le molte immagini esaminate, mostrano che la formazione di gruppi, anche, con oltre i 30 soggetti menzionati nell'articolo come massimo, sono tutt'altro che occasionali.
Nota: nella serie sottoesposta (3 foto) gli esemplari di Crossoptilon c. crossptilon mostrano quello che dovrebbe essere la colorazione tipo per questa specie nonostante il grigiastro su spalle e ali sia poco evidente. Da considerare anche una certa variabilità fenotipica individuale.
Foto da: 1 www.inaturalist.org Autore: Lin Sun Fong - 2-3 www.birdnet.cn ( località indicata dal fotografo: 1 prefettura autonoma tibetana di Garzê, Sichuan 10-10-2024 Cina - 2-3 prefettura autonoma tibetana di Garzê, Sichuan 25-06-2018 e 07-02-2020 Cina )
1 - Crossoptilon c. crossoptilon 2 - Crossoptilon c. crossoptilon 3 - Crossoptilon c. crossoptilon

.jpg)
.jpg)
Crossoptilon c. crossoptilon
Nota: le successive 4 immagini, nelle quali le favorevoli condizioni di luce mettono in risalto l'intesa sfumatura grigiastra che si riflette su gran parte della livrea, spiegano ancor meglio delle parole il dubbio che il Crossoptilon c. dolani possa rappresentare una mutazione più scura della forma nominale e di conseguenza anche l'origine del congenere Crossoptilon harmani.
Foto da: 1-2 wwwbirdnet.cn Autore: Xián tíng xìnbùYes ( località indicata dal fotografo: 1-2 solo provincia del Sichuan 04-02-2025 e 13-02-25 Cina )
1 - Crossoptilon c. crossoptilon 2 - Crossoptilon c. crossoptilon
.jpg)
.jpg)
Crossoptilon c. crossoptilon
Foto da: 1-2 wwwbirdnet.cn Autore: Xián tíng xìnbùYes ( località indicata dal fotografo: 1-2 solo provincia del Sichuan 30-12-2024 e 26-01-25 Cina )
1 - Crossoptilon c. crossoptilon 2 - Crossoptilon c. crossoptilon
.jpg)
.jpg)
Crossoptilon c. drouyini ( Fagiano orecchiuto bianco di Drouyin ) ........Verreaux Jules Pierre ......1868
Descrizione originale in:
NOUVELLES ARCHIVES DU MUSÉUM D'HISTOIRE NATURELLE. DE PARIS TOME QUATRIÈME 1868
DESCRIPTION de quelques NOUVELLES ESPÈCES D'OISEAUX Par M. J. VERREAUX (Aide-Naturaliste au Museum)
Crossoptilon drouynii - Milne Edwards (Pl. III.)
Candidum leviter supra cinerascens: rachi remigum nigra ; vertice plumis nigerrimis. bre-vibus, sericeis vestito ; cauda longissima; rectricibus basi albis, medio metallice viridibus, apice cceruleo-cupreis; facie rubro-carminea, papillosa, rostro pedibusque rubidis.
Bianco, leggermente grigiastro nella parte superiore: il rachide dei rematori è nero; la parte superiore è ricoperta di piume nerissime. Corto, setoso; la coda è molto lunga; le penne del dorso sono bianche alla base, verde metallico al centro e rosso ramato all'estremità; la faccia è rosso-carminio, papillosa, il becco e le zampe sono rossi.
Colore generale bianco, sfumato di grigiastro superiormente, soprattutto sulle ali e principalmente alla base delle remiganti primarie, dove questo colore si ritrova mescolato al nerastro che colora parte delle barbe interne. Il rachide di tutte le remiganti primarie e secondarie è nero lucido; la sommità del capo è nera, con piume corte, fitte e vellutate, sulle quali si evince un leggero bordo grigio argenteo, più distinto sul bordo che al centro; le penne della regione parotica, scomposte come nelle specie già note e soprattutto in quella considerata, per il momento, come Phasianus auritus di Pallas, sono lunghe circa quattro centimetri e formano su ciascun lato del capo un lobo auricolare rivolto verso l'alto. La coda, molto lunga e ampia, è composta da venti larghe timoniere con barbe meno scomposte che in quest'ultima, ad eccezione delle due mediane, le cui barbe sono strette come in C. auritum, tranne verso l'estremità che è larga come nelle altre e leggermente curva; una forma che si ritrova nelle cinque specie successive su ciascun lato. Queste timoniere, il cui rachide è nero, si presentano in grigio bianco dalla base, mescolato a nerastro, ma tutte ampiamente terminate in un bellissimo blu acciaio, con la parte mediana che mostra del verde violaceo e riflessi ramati. Faccia nuda, papillosa e rosso carminio, come nelle altre specie; becco rossastro, tarsi più scuri; unghie cornee.
Lunghezza totale…………………………………………………………....94 cm. >> mill.
— del becco dalla sua base superiore…………………………......3 - 6 -
— del tarso………………………………………………………………….….....9 - 6 -
— del dito esterno del piede, dove si rompe l'unghia……..5 - 6 -
— del medio……………………………………………………………………....7 - 9 -
— dell’esterno…………………………………………………………………....5 - >> -
— del pollice……………………………………………………………………....4 - >> -
— dell’ala chiusa……………………………………………………………….45 - >> -
— della coda seguendo la curvatura delle penne…...........30 - >> -
Questa magnifica specie, inviata al signor Soubeiran dal signor Dabry, console francese a Han-Keou, e donata al Museo di Parigi dal signor Drouyn de Lhuys, fu presentata all'Accademia delle Scienze dall'illustre professore di zoologia al Museo di Storia Naturale, il signor Milne Edwards, nella riunione del 16 aprile 1868. Dimostra una grande somiglianza con il Crossoptilon thibetanum, descritto dal signor Hodgson nell'Asialie Journal, vol. VII, p. 11, e raffigurato dal signor G. R. Gray nel suo “Genera of Birds”.
In generale, il suo piumaggio è altrettanto disgregato come in quest'ultimo, ma si distingue a prima vista per la colorazione delle penne remiganti, che sono marroni nel C. thibetanum e bianco-grigiastre nella nostra nuova specie. Ma ciò che presenta ancora maggiori differenze sono le timoniere esterne, che non solo non sono nere, ma mancano anche delle caratteristiche macchie bianche della specie indicata. Inoltre, le timoniere mediane non presentano né la larghezza né la brillantezza verde-oro della specie descritta dal signor Hodgson, il cui unico esemplare è da tempo nella collezione del British Museum.
Anche questa forma proviene dal Tibet, nella zona chiamata Mou-Pin; purtroppo, non abbiamo ancora informazioni precise sulle sue abitudini. Tutto ciò che possiamo affermare è che, a giudicare dalle altre specie conosciute, le femmine differiscono dai maschi solo per la mancanza di speroni, una caratteristica che separa questo genere dai veri fagiani, le cui femmine sono sempre dissimili, non solo nella colorazione del piumaggio, ma anche nelle dimensioni. Ci siamo resi doverosi e lieti di mantenere qui il nome di C. Drouynii, con cui il professor Milne Edwards lo aveva presentato all'Istituto, e come omaggio agli sforzi che il signor Drouyn de Lhuys, il degno presidente della Società per l'Acclimatazione, non manca mai di compiere, ogni volta che può rendere servizi alla scienza.
Caratteristiche subspecifiche: simile ai congenere "bianchi", misura mediamente da 95 a 105 cm., con un peso compreso tra 1400 e 2200 g. Si distingue da questi ultimi per la livrea quasi del tutto bianca eccetto la sommità del capo nera, pochissimo grigio sulle ali, limitato generalmente ai vessilli interni delle primarie ed una quantità variabile di bianco o grigio, solitamente mostrato da 4-o 5 delle timoniere più esterne, nella coda verde bronzo.
Foto da: 1-2-3 www.birdnet.cn Autori: 1 Qīnghǎi lù - 2 Yúshù jié - 3 Aichimù ( localitla indicate dai fotografi: 1 solo Qinghai 2020 Cina - 2 Kanda Mountain, prefettura di Yushu, Qinghai, 29-06-2021 Cina - 3 Prefettura autonoma tibetana di Hainan, Qinghai 04-07-2021 Cina ) - Nota: quest'ultima località parrebbe un errore quand'anche le foto fossero state scattate nell'estremo sud della prefettura, in quanto risulterebbe comunque, perlomeno in base alle informazioni attuali, un poco troppo a nord per questa forma. Se così fosse, in dette aree potrebbe interagire con il congenere Crossoptilo auritum.
1 - Crossoptilon c. drouyni 2 - Crossoptilon c. drouyni 3 - Crossoptilon c. drouyni

.jpg)
.jpg)
Crossoptilon c. dolani ( Fagiano orecchiuto bianco di Dolan )........Meyer de Schauensee.......1837
Nota descrittiva originale da : ZOOLOGICAL RESULTS OF THE SECOND DOLAN EXPEDITION TO WESTERN CHINA AND EASTERN TIBET, 1934-1936.
BY Brooke Dolan, II. Research Associate, Academy of Natural Sciences of Philadelphia
"...Il 22 maggio 1935 melle valli boscose intorno a Jyekundo (Tibet), Schafer catturò una serie di fagiani orecchiuti di una specie intermedia tra Crossoptilon c. crossoptilon e harmani, che Rodolphe de Schauensee mi ha fatto l'onore di chiamare Crossoptilon c. dolani. Questa forma è abbastanza simile al Crossoptilon harmani, ma di colore grigio più chiaro. Il suo piumaggio è ruvido al tatto, meno setoso rispetto a quello delle altre forme..."
"..Cento sfumature di grigio..", così potrebbe essere riassunto il fenotipo mostrato da questa forma nelle molte immagini esaminate. Tutto lascia ad intendere che potesse trattarsi, originariamente, di una mutazione della forma nominale, o più verosimilmente, perlomeno per quel che riguarda la sua "storia" ecologica contemporanea, un ibrido probabilmente prodotto da un primo incrocio tra Crossoptilon harmani e Crossoptilon drouyni e/o crossoptilon con successivi re-incroci in piena autonomia riproduttiva tra ibridi stessi o tra questi ultimi e le forme genitoriali determinando la moltitudine di varietà cromatiche osservabili, spesso, all'interno dello stesso gruppo con soggetto simili, se non identici, a drouyni o harmani, oppure altri in tutte le gradazioni intermedie di colore. Quindi, relativamente a quanto appena menzionato, ed al fatto che non esiste una vera separazione tra le gamme, risulta difficile considerare specificatamente distinto il Crossoptilon dolani.
Foto da: 1 www.ebird.org Autore: 1 Argrit Boonsanguan - 2 Autore: Wu Jiawei - 3 Autore: Xavier Rufray ( località indicate dai fotografi: 1 gradi decimali (32.828, 97.191) = contea di Yushu, prefettura di Yushu, Qinghai 01-06-2025 Cina - 2 distretto di Angsai, contea di Yushu, prefettura di Yushu, Qinghai <no data> Cina - 3 sottodistretto di Gyegu a Yushu City, prefettura di Yushu, Qinghai 13-05-2019 Cina )
1 - Crossoptilon c. dolani 2 - Crossoptilon c. dolani 3 - Crossoptilon c. dolani
%2C%20Qinghai%2001-06-25%20-%20Argrit%20Boonsanguan%20ebird%20org_.jpg)


Crossoptilon c. dolani
Foto da: 1 www.ebird.org Autore: Ma Yan Bryan - 2-3 www.ebird.org Autore: Robert Tizard ( località indicate dai fotografi: 1Kandashan, contea di Nangqian, prefettura di Yushu, Qinghai 07-06-2024 Cina - 2-3 Jiangxi Forest, Qinghai 02-07-2013 Cina )
1 - Crossoptilon c. dolani 2 - Crossoptilon c. dolani 3 - Crossoptilon c. dolani



Crossoptilon c. dolani
Come ben si evince dalla località indicata, il maschio delle foto sottoposte si trova in piena area drouyni/dolani, tuttavia mostra una colorazione solo leggermente più chiara rispetto al congenere harmani ma, soprattutto, non presenta le timoniere più esterne bianche come nei "tipici" dolani e drouyni.
Foto da: 1 www.ebird.org Autore: Robert Tizard ( località indicata dal fotografo in gradi decimali: 32.7007,95.5880 che corrispondo su Google Map a = PH2Q+75H Angsaixiang, contea di Zadoi, prefettura di Yushu, provincia del Qinghai 02-01-2020 Cina )
1 - Crossoptilon c. dolani 2 - Crossoptilon c. dolani 3 - Crossoptilon c. dolani


%2C%20Qinghai%2C%20China%2002-01-20%20-%20Robert%20Tizard%20ebird%20org%20%204.jpg)
Crossoptilon c. dolani
Purtroppo per la prima immagini non era indicata nessuna località. Il fenotipo parrebbe quello del Crossoptilon harmani, ma le timoniere esterne bianche suggeriscono la natura ibrida di questo soggetto, quindi attribuibile al Crossoptilon dolani. Non meno interessante l'esemplare della seconda foto ed in particolare la località menzionata ubicata nel nord del Tibet, area nella quale, secondo quanto menzionato nella letteratura, non dovrebbe trovarsi il Crossoptilon harmani. Condizionale d’obbligo in quanto, per produrre un ibrido serve l’incrocio tra due forme che nello specifico dovrebbero essere il Crossoptilon harmani e drouyni ed inequivocabili immagini confermano la presenza di harmani nel nord della prefettura di i Nyingchi o di Linzhi, quindi non lontano dai luoghi dove sono stati fotografati altri esemplari con piumaggio definibile intermedio.
Foto da: 1 www.idealera.com Autore: Feiyu - 2 www.birdnet.cn Autore: Wu Hanhui ( lovaità indicate dei fotografi: 1 nessuna info - 2 Prefettura di Nagqu, Tibet 03-09-2020 Cina )
1 - Crossoptilon c. dolani ? 2 - Crossoptilon c. dolani

.jpg)
Crossoptilon c. dolani
Visivamente, e di primo acchito, i due esemplari delle immagini sottoposte, potrebbero essere scambiati per dei Crossoptilon dolani che, a mio parere, di per se non dovrebbe essere considerato specificatamente distinto, in quanto lo sembra né più né meno di altre forme apparentemente più uniformi nel fenotipo (vedi leucurum) e senza un significativo intervallo tra le varie distribuzioni che, tra l'altro, il più delle volte si accavallano. Tuttavia l'ampia barratura bianca sulle ali e la coda pressochè solo di questo colore, ne suggeriscono la natura ibrida
Foto da: 1-2 www.birdnet.cn Autori: 1 良好的光影摄影 - 2 Gehua N.N ( località indicata dai fotografi: 1-2 prefettura autonoma tibetana di Yushu, Qinghai 05-03-2020 e 10-03-2023 )
1 - Crossoptilon c. dolani ? 2 - Crossoptilon c. dolani ?
.jpg)
.jpg)
Crossoptilon c. lichiangense ( Fagiano orecchiuto bianco di Lijiang )........Delacour J.T. ........1945
Descrizione originale di delacour del 1945 già sopra pubblicata: "..differisce dal Crossoptilon c. crossoptilon, al quale assomiglia di più, per le ali in grigio più chiaro; vessilli esterni delle primarie grigio chiaro; secondarie e terziarie grigio chiaro nella loro parte esposta, così come le copritrici alari. Le quattro paia esterne di timoniere presentano nella maggior parte dei casi un distinto bordo esterno grigio-biancastro, solitamente assente in Crossoptilon c. crossoptilon, e più grigio sulla metà basale di tutte le timoniere; sfumatura grigia del mantello mediamente più debole. I maschi adulti sono più chiari delle femmine e degli esemplari giovani, come in tutte le altre forme. Iride giallo oro pallido; cera grigia; becco rosa carnoso, giallastro verso la base; zampe e piedi scarlatti, artigli marrone-corno; pelle intorno agli occhi di un cremisi intenso e intenso.
Tipo 5, n. 543127, American Museum of Natural History, New York, coll., di C. Forrest, fianco orientale della catena montuosa del Lichiang, Yunnan nord-occidentale, altitudine 14.000 piedi, 10 ottobre 1922. Ali 327; coda 520; tarso 95; culmen (dalla narice) 36 mm. Otto esemplari esaminati; uno presenta 21 timoniere..."
Commento personale: se il Crossoptilon c. dolani, a fronte di valide disquisizioni, parrebbe da considerare un ibrido, ancor di più lo dovrebbe essere il Crossoptilon c. lichiangense la cui distinzione specifica sembrerebbe azzardata. Con un fenotipo che si distingue dai conspecifici di riferimento solo per essere leggermente più chiaro in alcune parti del piumaggio, ed una distribuzione molto approssimativa estesa tra i congenere Crossoptilon c. crossoptilon e drouyni, il dubitare ne divine una logica conseguenza. Del resto, se bastano i caratteri sopra esposti come determinante subspecifica, non è comprensibile perché il Crossoptilon leucurum sia considerato non sufficientemente distinto
Nota: come ben si evince dalle indicazioni di località, le foto sono state scattate un poco più a nord della gamma stabilita per questa forma, che comunque in base alle conoscenze attuali andrebbe del tutto rivista, tuttavia, la colorazione mostrata in queste immagini parrebbe identificabile con la descrizione sopra esposta. Come per gli Ithaginis, anche in questo caso a causa di informazioni sbagliate, e senza una precisa figurazione, avevo presunto che il fenotipo di questa forma fosse diverso. In realtà, da quanto esposto da Delacour, traspare un piumaggio che sembra intermedio tra il Crossoptilon c. crossoptilon e drouyni. A tale proposito occorre anche ricordare che la gamma del Crossoptilon c. lichiangense si trova circoscritta da quelle dei due precedenti congenere citati (vedere mappa di Delacour sopra esposta).
%2007-08-18.jpg)
Crossoptilon c. lichiangense: www.naturepl.com Autore: Staffan Wistrand (Baima Snow Mountain Nature Reserve, Yunnan 07-08-2018 Cina )
Foto da: 1-3 www.naturepl.com Autore: Staffan Wistrand - 2 www.douyin.com by: @Deqin County Media ( località indicate dai fotografi: 1-3 Baima Snow Mountain Nature Reserve, Yunnan 07-08-2018 Cina - 2 Baima Snow Mountain, Yunnan 02-05-2019 Cina )
1 - Crossoptilon c. lichiangense 2 - Crossoptilon c. lichiangense 3 - Crossoptilon c. lichiangense
%2007-08-18.jpg)

%2007-08-18.jpg)
Crossoptilon mantchuricum
Da "MONOGRAPH of the PHEASANTS" 1918 di William Beebe Vol. 1
Sinonimi:
Crossoptilon tibetanum Lamprey (nec Hodgs.), Proc. Zool. Soc., 1862, p. 221.
Crossoptilon auritum sive (= ovvero) mantchuricum Swinhoe, Proc. Zool. Soc., 1862, p. 286; Swinhoe, Proc. Zool. Soc., 1863, p. 306 [Manchuria].
Crossoptilon auritum Sclater, List of Phas., 1863, p. 6, pl. 5 [Manchuria]; Milne-Edward, N. Arch. Mus. Bull., I. 1865, p. 12, pl. 1, figs. 1 and 2 [E. of Pekin]; Sclater, Proc. Zool. Soc., 1866, p. 418; Saurin, Proc. Zool. Soc., 1866, p. 437 [Mts. N.W. of Pekin]; David, N. Arch. Mus, Bull, III. 1867, p. 37 [San-Yu]; Bartlett, Proc. Zool. Soc., 1868, p. 115; Saint-Hilaire, Bull..Soc. d’Acclim., (2), VII. 1870, p. 135 [breeding in captivity]; Gould, Birds Asia, VII. 1870, pl. 22.
Crossoptilon mantchuricum Swinhoe, Proc. Zool. Soc., 1863, p. 306; Newton, Ibis, 1865, p. 361 [Pekin] Gray, List Gallinae Brit. Mus., 1867, p. 31 ; id. Hand-list Birds, II. 1870, p. 259; David, N. Arch. Mus. Bull., VII. 1871, p. 11-[Pekin];: Swinhoe, Proc. Zool. Sec., 1871, p. 390; Sclater, Proc. Zool. Soc, 1871, p. 495 ; Mairet, Bull. Soc. d’Acclim., 1871, p. 594; Elliot, Mon. Phas., I. 1872, pl. 16: Cornély, Bull. Soc. d’Acclim., 1874, p. 168; David and Oustalet, Ois. Chine, 1877, p. 405, pl. 106 [Mts. of Pechili]; Sclater, Proc. Zool. Soc., 1879, p. 118, pl. VIII. fig. 5; Garrod, Proc. Zool. Soc., 1879, p. 373; Sclater, List Animals in Zool. Soc. Gardens, 1883, p. 477; Evans, Ibis, 1891, p. 76; Grant, Cat. Game-birds, XXII, 1893, p. 294; Tegetmeier, Pheasants, 1904, p. 228; Rothschild, Bull. Brit. Orn. Club, XIV. 1904, p. 58; Mitchell, Proc. Zool. Soc., 1911, p. 521; Beebe, Zoologica, I. No. 15, 1914, p. 275; Baker, Jour. Bomb. Nat. His. Soc., XXIV. 1916, p. 636.
Crossoptilon Lavison, Bull. Soc. d’Acclim., XI. 1854, p. 718 [first arrival at the Jardin des Plantes].
Crossoptilon manchuricum Seebohm, Bull. Brit. Orn. Club, I. 1892, p. 18 ; Grant, Hand-book Game-birds, I. 1895, p. 254; Goodchild, Bird Notes, IV. 1905, p. 65; Finn, Game-birds India and Asia, IgII, p. 84.
Crossoptilum manchuricum Sharpe, Hand-list Birds, I. 1899, p. 35 ; Oates, Cat. Eggs Brit. Mus., I. 1901, p. 53; Lanning, Wild Life in China, 1911, p. 112.
Crossoptilum mantchuricum Dresser, Manual Palae. Birds, II. 1903, p. 672; Ingram, Ibis, 1909, p. 462
Storia "antica":
Nel 1862 Swinhoe espose la pelle di una femmina di fagiano orecchiuto bruno che gli era stata inviata da Tientsin (o Tianjin) dall'amico Dr. Lamprey. Dopo molte vaghe teorie e perifrasi, decise che non si trattava di Crossoptilon auritum, sebbene questa supposizione costituisse il titolo del suo articolo, e quindi gli attribuì l'appellativo mantchuricum.
Eccetto un decennio di confusione con il fagiano orecchiuto blu (Crossoptilon auritum), la sinonimia di questa specie è pressoché priva di errori.
Raffigurazione molto accurata di Crossoptilon mantchuricum del 1872 realizzata da J. Wolf e J. Smit per Daniel Giraud Elliot.

Maschio adulto. - L'intera sommità della testa è ricoperta da una fitta crescita di piume corte e vellutate, nere e ricurve, ampie zone facciali nude di colore scarlatto, ricoperte da piccole papille. Piume appena dietro le narici, il mento, la gola e le piume sotto la zona facciale nuda, di colore bianco crema; le copritrici auricolari, molto allungate e rigide, sono di un bianco argenteo scintillante. Queste ultime sono rivolte verso l'alto e si estendono oltre il piumaggio su ciascun lato della testa come due ciuffi obliqui o "orecchie". Occipite e collo sono di un nero lucido tutt'intorno. Le file anteriori di penne occipitali presentano aree mediane e basali biancastre, e la brusca linea di penne della corona ricurve mette in evidenza queste parti, dando origine a una linea trasversale molto stretta, più o meno distinta, di colore bianco opaco in corrispondenza di questa giunzione.
Sul piumaggio superiore, il collo nero sfuma gradualmente in marrone sul mantello inferiore e sulle ali, e tutte le parti visibili delle penne sono lasse e pelose a causa del brusco accorciamento delle barbule e della scomparsa dei barbicelli sull'intera metà distale del vessillo. Anche sulla porzione basale, questi ultimi sono pochi e conferiscono scarsa coesione alla membrana. Le copritrici alari hanno una consistenza piuttosto soda, ma anche le secondarie e le primarie sono molto più morbide e flessibili rispetto alla maggior parte dei fagiani. Le copritrici alari e le secondarie sono fortemente lucide di porpora, mentre le membrane interne delle primarie mostrano una lucentezza bluastra altrettanto intensa. La parte inferiore del dorso, Il groppone e le copritrici caudali superiori sono di un bianco opaco e argenteo; la prima area è piuttosto macchiata da tracce di marrone dorsale. Sulla superficie ventrale, il collo nero sfuma nel marrone del sottopiuma rimanente. I fianchi e le copritrici caudali inferiori sono di un marrone leggermente più chiaro rispetto alle penne circostanti.
La coda composta da 22 penne è il fiore all'occhiello di questo fagiano dal colore piuttosto cupo. È fortemente graduata, con le penne centrali lunghe due volte e mezzo quelle esterne. La porzione di gran lunga più grande della penna è bianco opaco, che si macchia di marrone verso la punta. La parte terminale è nero brunastro lucido con un intenso blu violaceo. Le due paia esterne sono palmate in modo piuttosto compatto, ma dalla terza verso l'interno, la membrana esterna mostra una disintegrazione delle barbe, che allo stesso tempo diventano allungate e curve. Questo fenomeno aumenta fino a quando, nella terza coppia a partire dalla coppia centrale, mentre la membrana interna è piuttosto solida, quella esterna a un livello corrispondente sulla penna mostra filamentosi. barbigli lunghi quattro volte più di quelli sul lato opposto dello stelo. Le due paia centrali di penne della coda nel maschio adulto sono quasi interamente filamentose, e le barbigli drappeggiano l'intera coda quando questa è chiusa. Anche in queste, tuttavia, è presente una piccola punta spatolata di colore viola compatto. Nella posizione abituale, le penne della coda sono tenute in due linee piuttosto verticali, e queste coppie centrali disintegrate sono quasi sempre notevolmente sollevate rispetto alle altre. Le estremità delle quattro paia interne sono nettamente depresse.
In una considerevole percentuale di uccelli maschi si riscontra una marcata tendenza alla comparsa del bianco nelle ali. Le basi delle primarie sono spesso di colore bianco chiaro o screziato, e le membrane esterne delle stesse penne sono talvolta vistosamente vermicolate o screziate dello stesso colore. Non è insolito che la maggior parte delle penne della superficie inferiore presenti una distinta fascia terminale bianca. Vi è anche una notevole variazione nella proporzione relativa di nero e marrone nel piumaggio, con il nero lucido che occasionalmente predomina e dà l'impressione di un fagiano nero corvino. Ciò è in parte, ma solo in piccola parte, dovuto all'usura, poiché il piumaggio appena prima della muta è leggermente più chiaro rispetto a quello dell'uccello appena muto.
Area facciale nuda, zampe, piedi e speroni scarlatti; becco color corno rossastro chiaro; artigli color corno; occhi marrone rossastro chiaro. Lunghezza: 1000; becco dalla narice: 27; ala: 30,6; coda: 54,4; tarso: 100, dito medio e artiglio: 7,5 mm. Speroni robusti, forti e di forma conica, da 1 a 1,3 cm.
Femmina adulta. - Non vi è una netta differenza di colore tra maschi e femmine, ma queste ultime sono notevolmente più piccole. Becco dalla narice: 2,6; ala: 29,0; coda: 52,0; tarso, 9,4; dito medio e artiglio, 7,3 cm. Gli speroni sono appena visibili nelle femmine, essendo scaglie affilate ma molto corte.
Come i loro congenere gli orecchiuti bruni differiscono per molti particolari essenziali dalle specie più comuni. Entrambi i sessi sono simili nel piumaggio e si distinguono solo per la presenza di speroni sulle zampe dei maschi. Le grandi dimensioni e le caratteristiche peculiari delle copritrici caudali li separano da qualsiasi gruppo affine. I primi esemplari vivi furono donati alla Zoological Society dal signor Dudley E. Saurin nel 1866; da allora altri sono stati importati e un numero considerevole è stato allevato in questo paese e sul continente.
Il Crossoptilon mantchuricum è più ragguardevole per la singolare disposizione del suo piumaggio che per la brillantezza della colorazione non avvicinandosi alle splendide tonalità dei veri fagiani o di molte altre forme strettamente affini. Il colore generale del corpo è marrone scuro; le penne della coda sono bianche con apici scuri. Deve il suo aspetto notevole soprattutto alle grandi dimensioni e peculiare caratteristica delle copritrici caudali, che iniziano dalla parte inferiore del dorso e in gran parte nascondono la coda vera e propria. Queste copritrici si presentano bianche con barbe separate, andando a formare un'elegante appendice al corpo. Zampe e piedi di colore rosso, di tipo rasoriale, con gli artigli leggermente ricurvi, come quelli dei polli comuni.
La testa è alquanto sorprendente nel suo aspetto generale; becco arcuato bianco carnoso pallido, in forte contrasto con la pelle rossa del viso, che a sua volta è messa in risalto dalle piume bianche che costituiscono le cosiddette "orecchie" che caratterizzano queste forme.
Il console Swinhoe scrisse: "Si trova sulle colline a nord di Pechino, in Manciuria, e viene portato in città durante l'inverno in gran numero, sia vivo che morto. È chiamato dagli indigeni Ho-ke. Le piume di questo uccello erano anticamente indossate dai guerrieri tartari. Non ho mai visto la specie allo stato selvatico".
Da www.chinadaily.com.cn - Sunto dell'articolo di: Zhu Xingxin a Taiyuan e Zhou Huiying - 05-12- 2024
“…La popolazione di fagiani orecchiuti bruni, specie protetta di primo livello in Cina, è in costante aumento ed il suo areale si sta espandendo, affermano gli esperti durante un recente evento commemorativo a Taiyuan, nella provincia dello Shanxi.
Questa forma unica si trova principalmente nelle province cinesi dello Shanxi, Shaanxi. Nel 1984, il governo dello Shanxi ha designato il fagiano come uccello simbolo della provincia.
"Negli anni '80 la specie era sull'orlo dell'estinzione. Dopo oltre quattro decenni di impegno costante, sono stati compiuti progressi significativi nella conservazione di questo fagiano.
Misure come il rafforzamento dell’informazione ed educazione, protezione dell'habitat, ricerca scientifica, allevamento di soccorso, trasferimento sperimentale e monitoraggio ed il controllo delle malattie della fauna selvatica hanno migliorato efficacemente le condizioni di sopravvivenza dei fagiani orecchiuti bruni".
Da un'indagine è emerso che i fagiani dalle orecchie marroni nello Shanxi ammontano a circa 19.000 esemplari, con un'area di distribuzione che indicativamente ricopre un area di 12.000 chilometri quadrati..."
Crossoptilon mantchuricum - www.shanghaibirding.com Autore: Craig Brelsford ( Tempio di Xuanzhong ai piedi dei Monti Lüliang, Shanxi 30-08-2020 Cina )

Crossoptilon mantchuricum
Foto da: 1 www.news.cgtn.com - 2 www.gamebirds.me Autore: Di Dev - 3 www.xinhuanet.com Autore: Mei Yongcun ( località indicata dal fotografo: 1 Linfen City, contea di Hongdong Shanxi 10-11-2024 Cina - 2-3 Monte Shibi, contea di Jiaocheng, Taiyuan, Shanxi 23-12-2012 e 12-10-2018 Cina )
1 - Crossoptilon mantchuricum 2 - Crossoptilon mantchuricum 3 - Crossoptilon mantchuricum



Crossoptilon mantchuricum
Sottoposte alcune immagini di quella che dovrebbe essere una coppia di Crossoptilon mantchuricum con prole.
Foto da: 1-2-3 www.news.cgtn.com ( località indicata dal fotografo: 1-2-3 Qiliyu Scenic Area, contea di Huozhou, Shanxi 15-06-2024 Cina )
1 - Crossoptilon mantchuricum 2 - Crossoptilon mantchuricum 3 - Crossoptilon mantchuricum



Crossoptilon harmani
Da: The IBIS - QUARTERLY JOURNAL OF ORNITHOLOGY.
EDITED BY OSBERT SALVIN; M.A., F.R.S., AND PHILIP LUTLEY SCLATER, M.A., Ph.D., F:R.S., SECRETARY TO THE ZOOLOGICAL SOCIETY OF LONDON.
VOL. V...1881, FOURTH SERIES.
CROSSOPTILON HARMANI, sp. nov. Di: Elwes, H. J. 1881 ( descrizione originale )
Illustrazione di Crossoptilo harmani allegata alla descrizione originale del 1881 di J. G. Keulemans.

"...Becco color corno sfumato di rosso, lungo 2,54 cm dall'apertura e profondo 1,9 cm alla narice. Occhi e uno spazio intorno all'occhio, lunghi 5,08 cm e profondi 2,5 cm, nudi, rossi. La sommità della testa è ricoperta da corte piume blu-nere vellutate. Una fascia bianca si trova sull'occipite, sul mento e al centro della gola, per uno spazio di circa 12,7 cm dal becco. Copritrici del collo prodotte, lunghe circa 5 cm, bianche. Il resto del collo, il dorso, le copritrici alari, il petto, i fianchi e le copritrici inferiori della coda sono bluastri ardesia scura. Copritrici della coda superiori sono lunghe, piuttosto più grigie del dorso. Centro del ventre bianco. Primarie e secondarie bluastre ardesia scura, con riflessi violacei. Ali lunghe 30,48 cm. Coda composta da venti penne graduate, la coppia centrale lunga circa 45,72 cm, la coppia laterale lunga circa 22,86 cm, viola bluastro, con riflessi viola e verdi sulle quattro o cinque paia centrali. Tarsi robusti, lunghi 10,16 cm, con speroni robusti. Dito medio, con artiglio, lungo 7,5 cm. Zampe e piedi rosso vermiglio.
Località tipo:- Tibet orientale, circa 240 km a est di Lhasa (Harman).
Questa forma assomiglia alla raffigurazione di C. auritum, Pall., riportata nel "Phasianide" di Elliot, per la colorazione generale e le macchie. Può, tuttavia, essere facilmente distinta dalla coda, che non ha il bianco al centro delle penne laterali ed è di colore completamente diverso. Il tipo di C. auritum, secondo Elliot, è andato perduto; tuttavia, Pallas lo descrive con diciotto remiganti. La tavola del signor Elliot è tratta da esemplari raccolti dall'Abbé Armand David nelle province di Shensi e Kokonor, e originariamente descritti come C. cerulescens, David ('Comptes Rendus, lxx. p. 538, 1870). La figura mostra venti penne della coda, come nel caso del mio uccello; ma che la specie raccolta da David sia effettivamente C. auritum, Pall., o no, il mio uccello è chiaramente distinto da entrambi, per quanto si possa accertare senza vedere gli esemplari stessi.
Per questa splendida specie sono debitore al Tenente Harman, R.E., che si è distinto come geometra ed esploratore dell'Himalaya orientale, in particolare nel Sikkim, dove ha lavorato per alcuni anni. Quando ero a Darjeeling, lo scorso dicembre, ho visto la pelle di quello che ho subito riconosciuto come un nuovo Crossoptilon appesa alla parete della sua stanza. Sfortunatamente non era mai stata adeguatamente conservata, ed era in uno stato così terribilmente tarlato che i resti, che mi ha gentilmente donato e che ora si trovano al British Museum, non meritano di essere conservati. Si sono tuttavia dimostrati sufficienti al signor Keulemans per realizzare un disegno molto accurato, il cui unico difetto è che le copritrici auricolari non sembrano nell'esemplare così fortemente sviluppate come nella figura. La pelle fu portata al signor Harman da uno dei suoi periti indigeni, il quale disse di averla procurata a 150 miglia a est di Lhasa, a un'altitudine di circa 6000 piedi, dove veniva trovata in branchi durante l'inverno.
Questa parte del Tibet non è mai stata visitata da nessun europeo né da nessuno dei cacciatori indigeni del defunto signor Mandelli e, avendo, come riportato, un clima molto più mite e una vegetazione più rigogliosa rispetto alle zone occidentali del Tibet, ci si può aspettare che produca un certo numero di specie notevoli e, finora, sconosciute.
Questo rende la quinta specie del genere, o, se C. tibetanum, Hodgs., e C. drouyni, Verr., dovessero rivelarsi identiche, la quarta specie nota; e sebbene sia probabile che, come nel genere Phasianus, le razze o specie locali di Crossoptilon finiranno per fondersi impercettibilmente l'una nell'altra, tuttavia, per quanto ne sappiamo attualmente, non vi è alcuna difficoltà a distinguerle..."
Crossoptilon harmani ( fagiano orecchiuto di Harman )........Elwes, H. J. ......1881
Foto da: 1-2-3 www.birdnet.cn Autore: Xián tíng xìnbùYes ( località indicata dal fotografo: 1-2-3 solo provincia del Tibet <1-2> 24-07-2023 - 3 22-01-2023 )
1 - Crossoptilon harmani 2 - Crossoptilon harmani 3 - Crossoptilon harmani

.jpg)
.jpg)
Interessantissimo lavoro di Stuart Baker, che non a caso reputo uno dei migliori autori del passato al quale fare riferimento, tratto da:
JOURNAL of the BOMBAY NATURAL HISTORY SOCIETY - June 1916 VoL. XXIV. No. 4 THE GAME BIRDS OF INDIA, BURMA AND CEYLON By E. C. Stuart Baker, F.L.S., F.Z.S., M.B.0.U. PART XIX.
Phasianidae
Genus—CROSSOPTILON.
Il genere Crossoptilon comprende le forme comunemente note come "fagiani orecchiuti" a causa delle loro copritrici auricolari talmente prolungate da sporgere ben sopra alla testa come due piccole corna. Il loro colore varia dal bianco quasi puro al grigio ardesia scuro o marrone cinereo, e nell'aspetto generale sono forse i più simili ai fagiani del genere Gennaeus, ma differiscono molto per le caratteristiche della coda, che è molto larga alla base, meno compressa e con i vessilli molto larghi, morbidi e decomposti.
Le diverse specie presentano da 20 a 24 penne della coda. Il gruppo comprendente tibetanum e drouyni mostra 20 timoniere, manchuricum 22 e auritum 24. Crossoptilon harmani sembra averne solo 20, ma tutte le pelli esaminate erano in piena muta. Le ali sono arrotondate, nella tipica forma dei fagiani, con la quinta e sesta primaria più lunghe mentre la prima è relativamente corta; i lati della testa sono nudi, rossi e ricoperti da sottili papille. I maschi presentano speroni corti e smussati sui tarsi, assenti nelle femmine.
I sessi sono apparentemente simili nel colore, ma è possibile che le femmine mantengano la colorazione più scura del primo piumaggio più a lungo dei maschi.
Si trovano solo a grandi altitudini nelle catene montuose che si estendono dal Tibet alla Cina orientale.
Nella Hand-List of Birds, Sharpe ammette cinque specie in questo genere, tra cui Crossoptilon tibetanum, leucurum, manchuricum, auritum e harmani.
Attualmente, il numero di esemplari di questi splendidi fagiani disponibili per l'esame è davvero troppo esiguo per consentire una decisione definitiva sulla misura in cui si integrano, su quali siano da considerare specie valide e quelle come varianti o sottospecie geografiche.
Sembrano esistere tre forme ben definite: una, tibetanum, in cui il colore superiore è praticamente bianco puro o grigio chiaro, una seconda, auritum, in cui il colore è grigio scuro, e una terza, manchuricum, in cui il colore superiore è cinereo o marrone piuttosto che grigio. Anche tra questi, tuttavia, non è sempre facile definire dove finisce tibetanum e inizia auritum, poiché difficilmente due uccelli sono esattamente uguali, e la mancanza di dati precisi su molte delle poche pelli che siamo stati in grado di esaminare complica ulteriormente l'argomento.
Quando arriviamo alla definizione delle sottospecie, il compito diviene ancora più arduo. Abbiamo, in alcuni casi, pelli di esemplari catturati durante la stessa spedizione, assegnate a forme diverse, sebbene assolutamente nulla dimostri che non siano state tutte raccolte nello stesso luogo, e, ancora, di una specie, il leucurum, abbiamo due individui identificati come tipi maschile e femminile che sono molto meno simili di diversi altri esemplari a cui viene assegnato un rango specifico diverso.
Sembra tuttavia che, oltre alle differenze primarie tra le forme bianca e grigia, vi siano due varianti minori in entrambe. In ciascuna forma esiste una "variante" che non presenta bianco sulle penne della coda, e una seconda in cui è generalmente presente una certa quantità di bianco o grigio, e inoltre l'assenza o la presenza di questo bianco è accompagnata da altre caratteristiche abbastanza stabili, come il colore delle membrane esterne delle primarie.
Sembra inoltre ampiamente dimostrato che queste variazioni si riscontrano in determinate aree, ai confini delle quali gli uccelli incontrati presentano caratteristiche intermedie, una condizione che ci si aspetta naturalmente in sottospecie geografiche che non hanno ancora raggiunto lo status di specie vere e proprie.
Abbiamo quindi due forme di "tibetano", una con e l'altra senza basi bianche sulle penne della coda, la prima con vessilli grigio-marroni sulle primarie più esterne, e la seconda con vessilli bianchi o biancastri, note come Crossoptilon tibetanum tibetanum e tibetanum drouynii.
Similare il caso del Crossptilon auritum con due forme, auritum harmani senza bianco sulla coda e auritum auritum con basi bianche sulle penne della coda e un anello bianco molto più ampio attorno alla testa.
Mi accorgo di non riuscire a distinguere tra i drouynii di Verreaux e leucurum di Seebohm, anche se potrebbe essere possibile farlo quando avremo più materiale con le località e date esatte.
Gli esemplari del British Museum non sono stati esaminati e revisionati per molto tempo, e molti sembrano essere stati erroneamente denominati. Così, mentre due degli uccelli di Thorrold e Bang del Passo Sok in Tibet appartengono ai tipi di leucurum, un altro raccolto vicino al Passo Sok dal Duca d'Orléans è etichettato come tibetanum, e altri ancora provenienti dalla stessa spedizione sono classificati come leucurum. Inoltre, ci sono tre splendidi esemplari raccolti da Thorrold e Bang nello stesso luogo e periodo dei tipi di leucurum etichettati come tibetanum; presumibilmente sono stati etichettati così perché non mostrano il bianco sulla coda, ma i loro disegni laterali grigi e le primarie grigio chiaro o bianche li indicano come leucurum o drouynii.
Il tipo Verreaux di drouynii fu raccolto da Pére David a Moupin e nel Museo sono presenti esemplari di questa località, etichettati tibetanum, raccolti da quest'ultimo, che, come gli esemplari di Thorrold e Bangs, possono essere distinti a colpo d'occhio dai veri tibetanum provenienti da altre zone per il piumaggio più bianco, le basi della coda grigie anziché nere e, soprattutto, per le ali quasi completamente bianche con membrane esterne bianche sulle primarie.
Il bianco sulle penne della coda del leucurum (vel. drouynii) sembra variare enormemente. Il maschio tipo di leucurum presenta solo le penne terminali della coda, lunghe circa cinque o sette centimetri, insieme ai peduncoli e alle strette strisce sui stessi, nere; il resto della coda è bianco, con sfumature grigio scuro che sfumano nel nero. Il tipo femminile mostra pochissimo bianco sulla coda, e in alcuni altri esemplari non ce n'è affatto, ma in ogni uccello le basi delle penne laterali della coda, che abbiano o meno del bianco, sono grigie, non nero lucido come nel vero tibetanum.
Verreaux, nella sua definizione latina di drouynii, scrive: "rectricibus basi albis", inducendo a supporre che le basi delle penne della coda siano bianco puro, ma nella descrizione francese dice delle penne esterne della coda: "Le timoniere, il cui rachide è nero, con una parte delle loro basi bianco grigiastra". Sia la tavola di Elliott che quella di Verreaux raffigurano le basi delle penne della coda in grigio piuttosto scuro, che vira al nero nelle loro metà terminali.
Elliott nella sua Monografia del Phasianide sottolinea che il drouyni differisce dal tibetanum "in primo luogo, per i colori o le tinte primarie che sono brunastre nel tibetanum ma bianco-grigiastre nella nostra nuova specie. Ma la differenza più grande riguarda le penne esterne della coda che non sono nere e altresì prive delle macchie bianche così caratteristiche nelle altre specie. Vorremmo anche notare che le penne centrali della coda non sono altrettanto grandi, né possiedono il verde dorato brillante degli uccelli descritti da Hodgson".
Va ricordato che l'esemplare tipo descritto da Hodgson e nominato come tibetanum è un soggetto alquanto anomalo, con numerose grandi macchie o chiazze bianche sulle penne della coda, un modello che non ho visto in nessun altro esemplare.
Se, quindi, come mi sembra dimostrato senza ombra di dubbio, drouynii e leucurun costituiscono la stessa forma, quest'ultimo diventa sinonimo del primo.
Caratteri subspecifici:
A.— Colore superiore bianco o grigio chiaro. a. Assenza di bianco o grigio sulla coda; vessilli esterni delle primarie scuri... tibetanum tibetanum.
b. Coda; penne in parte bianche o grigie; vessilli esterni delle primarie bianchi o grigio molto pallido ... tibetanum drouynii.
B.— Colore grigio scuro nella parte superiore.
a. Nessun bianco sulla coda.... auritum, harmani. b. Piume della coda parzialmente bianche .. auritum auritum.
C.— Colore superiore brunastro, non grigio ... manchuricum.
La posizione di harmani è piuttosto difficile da definire. Nell'aspetto generale è molto simile all'auritum e potrebbe essere considerato come una sottospecie di quest'ultimo. Per contro, il numero di penne costituenti la coda sembrerebbe suggerire che sia più vicino al tibetanum.
Il gruppo tibetanum (leucurum) drouynii e harmani presenta 20 penne della coda, 24 auritum e manchuricum 22.
Il tipo di harmani presenta 19 penne ed è incompleto; il maschio e la femmina offerti al Museo dal Capitano Bailey sono in muta ma sembrano aver avuto 20 penne della coda; gli altri esemplari della bella serie ottenuta da quest'ultimo collezionista, ad eccezione di una pelle, sono ora semplicemente esemplari impagliati in una teca di vetro in una collezione privata, ma sono stati esaminati da me prima di essere montati e, sebbene nella maggior parte dei casi incompleti o in muta, sembravano non avere mai avuto più di 20 penne della coda. Se alla fine ciò dovesse rivelarsi invariabilmente il caso, harmani dovrà essere elevato al rango di specie e la chiave (caratteri subspecifici) di cui sopra dovrà essere modificata di conseguenza.
CROSSOPTILON TIBETANUM TIBETANUM Hodgson (Attuale Crossoptilon c. crossoptilon)
Phasianus (Crossoptilon) tibetanus.—Hodgson, Journ. As. Soc., ‘Beng. VII., p. 864, pl. 46 (1838), id., Ind. Rev. III., p. 593, pl. (1839). Crossoptilon tibetanum.—Hodgson in, Gray’s Zool. Misc. p. 85 (1844) ; Sclater, List Phasianide p. 6, pl. 4 (1863) ; David Vou. Arch. Mus. Bull, VII., p.11 (1871); Elliot, Mon. Phasianidee, 1, pl. 14 (1872); David and Oustalet, Ois. Chine, p. 407, pl. 107 (1877); Hume, Stray Feath., VII., p. 426 (1878); Scully, Stray Feath., VIIT., p. 343 (1879) ; Oustalet, Ann. Sci. Nat. (7), XII. p. 315 (1892) [part]; Ogilvie-Grant, Cat. Birds B. M., XXII., p. 298 (1893) ; Ogilvie-Grant, Hand-L. Game Birds, I., p. 252 (1895); Davies, Ibis (1901) p. 409; Bailey, Journ. Geog. Soc., XXXIX., pp. 339, 346 (1912) id Bom. Nat. His. Journ., XXII., p. 867 (1913); Finn, Indian Sporting Birds, p. 194 (1915). Crossoptilon auritum.—Gray, Genera B., III., p. 495, pl. CXXV. (1845). Crossoptilon tibetanum.—Hume and Marshall, Game-Birds, Ind. I, p. 115, pl. (1878) ; Seebhom, Ibis (1891) p. 378; Sharpe, HandL. I, p. 35 (1898); Oates, Eggs B. M., p. 53 (1901); Dresser, Manual Pal. B., p. 671 (1908).
Nomi vernacolari.—Chakgong, Chakai (Tibetan); Boté-Dafé (Nepal) ; Machi (Chinese).
Descrizione: Maschio e femmina adulti. - Sommità della testa ricoperta da piume corte, nere, arricciate e vellutate; penne alari marrone cenere, con le secondarie interne più grigio ardesia, leggermente iridescenti nelle parti blu visibili; le copritrici alari sfumano dal bianco, appena soffuse di grigio sulle copritrici più piccole, a grigio quasi scuro come sulle primarie e copritrici maggiori; coda bronzo porpora alla base, che sfuma dal verde metallico al blu-viola intenso sulla metà terminale. Il resto del piumaggio è bianco.
La purezza del bianco varia notevolmente da individuo a individuo; in alcuni è praticamente bianco puro, ma nella maggior parte dei casi è molto soffuso di grigio. In due esemplari il dorso è sufficientemente grigio da far sì che questa parte del piumaggio contrasti nettamente con il bianco puro della parte inferiore del dorso e groppone.
Colori delle parti molli. Maschio. - Iride giallo-rossastra, zampe rosso scarlatto brillante, artigli perlopiù cornei con apice più scuro; pelle nuda del viso rossa.
Femmina. — Lunghezza 80,01 cm. Iride gialla, becco rosato, zampe scarlatte, dita grigie. "Ba-la-to 3647 metri di altitudine" (F. M. Bailey). La pelle nuda della femmina è scarlatta, praticamente brillante come quella del maschio; dita rosso corneo, con estremità più scura.
Dimensioni generali: lunghezza da 76,2 a 88,9 cm.; becco anteriore circa 44,5 mm., e dalla bocca circa 50,8 mm.; tarso circa 88,9 mm.; coda da 457,2 a 559,0 mm.; ala da 292,1 mm., a 386,5 mm.
Con il materiale disponibile per il confronto, costituito in gran parte da esemplari asessuati, è impossibile dire se vi sia una differenza nelle dimensioni dei sessi, ma sembrerebbe di no.
Distribuzione: montagne della Cina occidentale e dell'estremo Tibet orientale. Sono ancora necessari dati più completi e accurati per stabilire dove questo uccello si incontra e si trasforma nella forma successiva, la drowynii.
A sud si trova comunque fino al Tongla, il passo all'inizio della valle dell'Irrawaddy, dove il Capitano F. M. Bailey lo trovò numeroso. Potrebbe benissimo essere rinvenuto in seguito nelle catene montuose più elevate del nord-est dello Yunnan 1*)
Nidificazione. — Non vi è alcuna documentazione sulla nidificazione di questo uccello in natura. Bailey lo ha trovato mentre nidificava a Tachienlu a metà maggio a circa 3600 metri, ma non è riuscito a trovare il suo nido.
Seebohm registra 5 uova di dimensioni variabili da 2°42" x 1,7" a 2,3" x 1,72". Il colore ricorda le uova di Perdix petrosa.
L'unico esemplare che possiedo nella mia collezione è uno acquistato per me da alcuni commercianti di piume tibetani che un tempo visitavano annualmente Darjiling per vendere la loro merce. Insieme all'uovo furono acquistate anche le pelli di due uccelli che si dice fossero i genitori. Si dice che questo esemplare sia stato catturato in un luogo chiamato Mizsim, nel nord-est del Tibet, tra il 1° e il 10 giugno 1892. L'aspetto delle pelli e delle uova al momento dell'acquisto confermava la data indicata. L'uovo è molto simile a quello di un comune pollo, ma ha una consistenza più liscia e satinata, con una grana più fine e fitta. È di colore grigio pallido e ha una forma ovale normale, misurando 2°2"' (= 55'4, mm) x 1:66" (= 42:1 mm). Non corrisponde nell'aspetto alle uova descritte da Seebohm, ma per consistenza, forma e dimensioni è simile a quelle di altre specie di fagiani dalle orecchie deposte in cattività. e differisce solo nel colore, essendo più camoscio e meno verde.
Al British Museum è presente una discreta serie di ego di questo fagiano, catturati da A. E. Pratt a Ta-tsien-la, composti da tre covate rispettivamente di tre, quattro e cinque uova. Tutte queste uova sono state catturate a maggio, la covata di cinque, evidentemente molto dura, quando è stata catturata il 18 di quel mese. Il colore di questi ego varia notevolmente: la covata di quattro è di un color pietra molto chiaro, quella di tre di un camoscio intenso e la terza covata è intermedia tra le due. Anche la consistenza differisce notevolmente: la prima covata menzionata ha una superficie molto fine e liscia, più simile a quella di un uovo d'anatra che a quella di un pollo, mentre la seconda ha la superficie ricoperta da innumerevoli minuscoli pori o fossette. Anche la terza covata è intermedia tra le altre due. Tutte hanno la venatura fitta, estremamente dura e con una notevole lucentezza. Tutte le uova potrebbero essere facilmente paragonate a quelle del pollo domestico.
La variazione in lunghezza è compresa tra 2°29" (58,2 mm) e 2:46"' (61,6 mm), e in larghezza tra 1°70" (43,2 mm) e 1,76" (44,7 mm), con una media delle dodici uova pari a 2°36" x 1,71" (59,9 x 43,4 mm).
La larghezza è particolarmente costante: le due covate più grandi contengono sette uova che misurano 43°2 mm e due che misurano 4,3°4 mm.
Abitudini generali. — La descrizione più completa delle abitudini di questo fagiano è quella fornita da Pére David, ma sebbene sia stata spesso citata come riferita al tibetanum, si riferisce senza dubbio, almeno in parte, al drouynii, il cui tipo è stato ottenuto da Pére David a Moupin.
Padre David scrive:
"Il fagiano di Hodgson vive nelle foreste di pini ad altitudini che variano dai 3.000 ai 3.600 metri sul livello del mare. Ha abitudini estremamente socievoli e si dice che se ne possano trovare 40 o 50 esemplari che si appollaiano in gruppi sui pini.
Si può trovare solo in alcune località boscose della Cina, sulle alte montagne del Sichuan occidentale, nei dintorni di Moupin e Ta-tsien-lou, dove la sua esistenza è protetta dal rispetto superstizioso degli indigeni. È un uccello molto gregario, che ama vivere in compagnia di molti dei suoi simili, anche quando è impegnato ad allevare i piccoli, e non si allontana molto dal luogo di riproduzione. Si nutre di foglie, radici, cereali e insetti. Fortunatamente per la sua incolumità, la carne di questo fagiano orecchiuto è solo moderatamente buona da mangiare, e i cacciatori preferiscono i fagiani più piccoli (Phasianus) come selvaggina, poiché sono più ampiamente distribuiti e più facili da reperire".
Davies in "Ibis" aggiunge qualche dettaglio in più. Scrive:
"Questi fagiani lepre erano comuni nelle stesse località dei fagiani rossi. Si trovavano in grandi stormi e correvano molto velocemente, volando a malapena, mentre erano così cauti che, sebbene ne abbia visti molti, non sono mai riuscito ad avvicinarmi abbastanza per sparargli. Il loro richiamo è un grido di corvo molto forte e aspro, che può essere udito per un miglio o due. Si mantengono ad altitudini elevate e spesso si trovano nella neve. Il punto più a sud in cui li ho visti è poco sopra i 28° di latitudine nord-occidentale di Chung-tien".
1*) Commento personale.- Probabilmente si tratta di "Lapsus calami" o errore di stampa dato che queste forme, nella provincia dello Yunnan, vivono nel nord ovest della stessa.
CROSSOPTILON TIBETANUM DROUYNI Verreaux
Crossoptilon drouyni.—Verreaux, Nouv. Arch. Mus. Bull., IV., p. 85, pl. ITI. (1868); Swinhoe, P.Z.S. (1871) p. 399; Elliott, Mon. Phasianidae 1, p. XVIIL.; pl. 15 (1872). Crossoptilon tebetanum.—Oustal., Ann. Sci. Nat. (7) XIL., p. 315 (1892) pt. Crossoptilon leucurum.—Seebohm, Bull. 1 BM OM Gee Dees oad 7 (1892), id. Ibis, 1893, p. 250; Ogilvie-Grant, Cat. Birds B.M., XXII., p. 294 (1898) ;id., Hand-L., Game Birds 1, p. 253 (1899). Crossoptilum leucurum.—Dresser, Manual Pal. Birds, p. 671 (1903).
Nomi vernacolari.—Chakgong, Chakai, (Tibetan); Boté-Daté, (Nepal) ; Machi (Chinese).
Descrizione.— Maschio e femmina adulti.— Differisce dal tibetanum per essere generalmente bianco molto più puro sul piumaggio superiore, specialmente sulle ali, mentre i vessilli esterni delle primarie sono bianco puro o grigio molto pallido anziché marrone o marrone chiaro in una tonalità piuttosto scura. Le basi delle penne caudali esterne sono spesso bianche; in alcuni esemplari, come nel tipo di leucurum, questo bianco si estende praticamente a tutte le penne caudali centrali e a tutte quelle esterne, in altri si estende solo a 4, 6 o 8 delle paia di penne esterne, mentre in alcuni non c'è praticamente bianco. In tutti gli esemplari di drouynii, tuttavia, le metà basali delle penne caudali, invece di essere nere lucide, come nel tibetanum, sono grigie.
I colori delle parti molli sono gli stessi del tibetanum.
Dimensioni come nel tibetanusm.
I tipi di leucurum furono raccolti da Thorrold e Bangs tra il Passo di Sokpo e Chiamdu, mentre il tipo di drouynii fu raccolto da Pére David a Moupin. Tuttavia, altri esemplari, raccolti in quest'ultimo luogo, si trovano al British Museum, oltre ad uno al Tring Museum, tutti denominati leucurum, mentre d'altra parte ci sono esemplari di leucurum raccolti dal Duca d'Orléans tra Sokpo e Chiamdu che, sebbene così denominati, risultano praticamente indistinguibili dal tipo di drouynii.
Questo è di per sé sufficiente a dimostrare che le due specie, leucurum e drouynii, sono la stessa forma, poiché non è possibile che due sottospecie vivano nello stesso habitat.
Il tipo di tibetanum è un uccello con macchie bianche sulla coda e sembrerebbe che questo bianco sia più un carattere individuale che subspecifico e i punti su cui fare affidamento per distinguere i drouyni dai tibetanum sono i colori primari e la loro relativa bianchezza, l'assenza o la presenza di grigio accanto al nero delle code.
Distribuzione: apparentemente a nord-ovest e ovest dell'area abitata dal Crossoptilon harmani, tra il passo di Sokpo e Chiamdu, e a Moupin. Non si conosce l'esatto punto in cui il Duca d'Orléans ottenne i suoi esemplari.
Nidificazione: nulla di noto.
Abitudini: le note di Padre David, come già riportato, si riferiscono a questa forma quanto, se non di più, rispetto alla precedente.
Sebbene l'esemplare specifico selezionato come tipo di femmina dalle serie raccolte da Thayer e Bangs sia quello descritto da Ogilvie-Grant nel suo Game Birds, va notato che non c'è motivo di pensare che la femmina differisca dal maschio più di quanto lo siano le femmine delle altre specie. La femmina tipo è sicuramente un uccello del tutto atipico, forse un esemplare giovane, e forse nemmeno una femmina. D'altro canto, il tipo maschile si rivelerà quasi certamente un esemplare eccezionalmente vecchio e bello, con una quantità anomala di bianco sulla coda.
CROSSOPTILON AURITUM AURITUM (Pallas)
Phasianus auritus.—Pall, Zoog. Rosso. Asiat. II., p. 86 (1811) Sclater, Ibis (1874) p. 170. Crossoptilon auritum.—Gray, Hand-L. Birds, II., p. 259 (1870) ; Swinhoe, Proc. Zool. Soc. (1871) p. 899; Sclater, Proc. Zool. Soc., (1871) p. 495 ; Elliot, Monog. Phasianide 1, pl. 17 (1872); Prjev., Mongolia, Strand Tangut, II., p. 121; pl. XX., fig. 1 (egg) id., in Rowley Orn. Misc. II., p. 420 (1877); David and Oustalet, Ois. Chine, p. 406, pl. 108 (1877); Prjev., Reisen in Tibet, p. 204 (1884) ; Pleske Bull., Acad. St. Petersb., XIII., p. 297 (1892); OgilvieGrant, Cat. Birds B. M., XXII., p. 295 (1898) ; id., Hand-L. Game Birds 1, p. 255 (1895); Berez. and Bianchi, Ptitz. Gan-su, pen 24i; - Styan, Ibis (1899) p. 298; Dresser, Manual Pal. B. p. 673 (1903) ; Parrot Filchner, Exped. China-Tibet, p. 132 (1908); (egg figured Gia lat jenzes Mongol. i., Strand Tanegut, i., p., 121, pl. xx., fig. 1). Crossoptilon caerulescens.—David, Ms. Milne-Edwards Comp. Rendu, LXX., p. 538 (1870); id., Ann. Mag. Nat. Hist. (4), V., p. 808 (1870) ; David, Nouv. Arch. Mus. Bull., VII., p. 11 (1871); Sclater, Ibis (1871) p. 370. Crossoptilum auritum.—Sharpe, Hand-L. vol. I., p. 35 (1898).
Nomi vernacolari.—Ushasty Fasan (Kansu); Hara-takia (Mongol); Shan-dgi (Chinese); Shariama (Tanquto).
Descrizione.— Maschio e femmina adulti.— Apice della testa, dalla fronte alla nuca, in nero vellutato; mento, gola, estremità anteriore del collo e copritrici auricolari allungate bianche. Parte superiore, grigio-blu ardesia chiaro; penne del collo vicino alle copritrici auricolari più scure, quasi nere, con le basi delle piume vicino alla corona bianche, con questo colore che si mostra come una sottile linea indistinta. Remiganti alari marroni, il resto dell'ala chiusa visibile come il dorso, le secondarie interne iridescenti blu-viola; parte inferiore come il dorso, ma leggermente cinereo sull'addome, copritrici sottocaudali e ano; coda con le lunghe penne centrali decomposte e quelle adiacenti grigio scuro con apici e bordi neri con iridescenze blu di Prussia; cinque o sei paia di penne esterne bianche con la terza terminale blu-nera lucida, la coppia adiacenti screziata di nero e bianco alla base.
Colori delle parti molli.— Iridi gialle e marroni, giallo-marroni o giallo-rossastre; pelle orbitale e palpebre scarlatte; zampe scarlatte o rosso scuro, speroni pallidi e corneo, con estremità più scura; becco rossastro e corneo.
Misure: lunghezza totale da 84 cm a 102 cm, a seconda di quanto è sviluppata la coda, che varia notevolmente in entrambi i sessi. Ali da 286,7., a 323,8 mm; becco anteriore circa 38,1 mm., e dalla bocca circa 44,4 mm; tarso circa 88,9 mm. Coda, da circa 457,2 mm a 596,9 mm. Quest'ultima è la più lunga che abbia mai visto, appartenente ad un esemplare asessuato conservato al Tring Museum. È un individuo anziano, senza alcun segno di speroni, ed è evidentemente una femmina.
Le copritrici auricolari di questa specie raggiungono i 63,5 mm., o anche di più, mentre una lunghezza di 50,8 mm., nelle forme bianche è davvero eccezionale.
Pulcinotto. — Superiore marrone nerastro molto scuro, con striature centrali color camoscio e barre indefinite in rossiccio e marrone; corona marrone scuro, non nera; sulla parte superiore del dorso compaiono le piume grigio ardesia; le penne delle ali sono screziate in marrone e rossiccio opaco, inferiormente color cenere sporco, il grigio ardesia dell'adulto è già visibile sul petto. Coda corta, di colore simile a quello dell'adulto.
Ala circa 228-6 mm.; copritrici auricolari ben sviluppate e lunghe oltre 25,4 mm.
Pelle orbitale rosso vivo; becco molto più scuro che nell'adulto, mascella quasi nera con bordi rossastri, mandibola rossastra e corneo. Le zampe sembrano essere state rosso corneo opaco, più luminose e rosse nella parte anteriore che in quella posteriore.
Distribuzione. — Catene montuose della Cina occidentale, Sichuan nord-occidentale, Gansu e Koko Nor.
Nidificazione.— Przhevalsky descrive le uova di questi fagiani rinvenute nei monti Gansu e Alaska (Alashan) come segue: "Le uova della specie attuale assomigliano nella forma a quelle del pollo da cortile, ma sono molto lisce e di un colore grigio oliva chiaro, senza macchie; diametro grande 5,08 cm, diametro piccolo da 4,06 a 4,14”.
Posseggo due uova di questa specie nella mia collezione, deposte da soggetti ospitati nel Giardino Zoologico e acquistate quando la collezione di Bartlett fu smantellata.
La loro forma è ovale piuttosto allungata, ma leggermente compressa verso l'estremità più piccola. La consistenza è molto fine e compatta, con una superficie lucida e straordinariamente liscia dalla colorazione grigio oliva chiaro, simili nell'aspetto generale più alle uova di anatra che a quelle di gallina. Un uovo, leggermente deforme, presenta anche numerosi piccoli noduli al centro, ma si tratta ovviamente di anomalie. Misurano 59,6 mm., x 4,6 mm., e 57,9 mm., x 39,3 mm.
Ci sono anche quattro uova deposte in cattività nella collezione del Museo di Storia Naturale; queste corrispondono bene a quelle della mia collezione, tuttavia, mentre due sono di colore molto simile alle mie, le altre due sono più tendenti al color pietra chiaro, senza la sfumatura olivastra.
Misurano tra 56,4 mm., e 58,4 mm., di lunghezza e tra 40,4 mm., e 41,4 mm., di larghezza; la media di tutte e otto le uova è 56,9 x 40,6 mm.
Przhevalsky fornisce la seguente descrizione delle abitudini del Fagiano orecchiuto di Pallas:
"..In entrambe località sopra menzionate, abitano le regioni boscose e montuose raggiungendo anche i 3.000 metri di altitudine sopra il livello del mare. Sono residenti e rimangono tutto l'anno in determinati luoghi. L'acqua non sembra essere determinante per questa specie, perlomeno non come per i congenere; ciò sembrerebbe evincersi dal fatto che sostano spesso in località sui monti Ala-Shan dove non si trova una goccia d'acqua.
In autunno e in inverno si riuniscono in piccoli stormi, probabilmente famiglie che, molto spesso, in primavera si dividono in coppie, quando i maschi iniziano a cantare emettendo a intervalli una nota forte e sgradevole che ricorda in qualche modo il richiamo del pavone. Solitamente al mattino, occasionalmente anche durante il giorno. All'inizio di maggio quasi tutte le femmine sono in cova. Il numero dele uova, secondo le dichiarazioni degli indigeni, varia da cinque a sette.
Dopo la stagione riproduttiva, i maschi iniziano subito la muta e riprendono il loro piumaggio fresco solo a ottobre. Generalmente le loro piume si consumano molto presto e gli uccelli indossano il piumaggio completo solo per un breve periodo in inverno e primavera.
Non siamo riusciti a trovare alcun cucciolo: ma il Tangut mi ha informato che i piccoli sono sempre accompagnati da entrambi i genitori. Sia i vecchi che i giovani corrono molto velocemente e sono molto abili nel nascondersi tra i fitti cespugli quando vengono inseguiti, e generalmente fanno affidamento più sulle zampe che sulle ali. Come la maggior parte della famiglia, questi uccelli amano scavare nel terreno in cerca di radici; e sembra che si nutrano principalmente di piante..".
CROSSOPTILON AURITUM HARMANI Elwes
Crossoptilon harmani.—Elwes, Ibis (1881) p. 399, pl. XIII; Ogilvie-Grant, Cat. Birds B. M. XXIT., p. 296 (1893); id., Hand-L. Game Birds I., p. 257 (1895); Dresser, Manual Pal. Birds, p. 673 (1903); Stuart Baker, Bull. B.O.C. XXXIII., p. 121 (1914); Elwes, Bull. B.O.C. XXXIII, p. 136 (1914); Bailey, Bom. Nat. His, Jour. XXIV,. pag. 77
Nomi vernacolari.—Cha-nea (Tibet).
Crossoptilon harmani in: JOURNAL of the BOMBAY NATURAL HISTORY SOCIETY - June 1916 VoL. XXIV. No. 4 THE GAME BIRDS OF INDIA, BURMA AND CEYLON - By E. C. Stuart Baker, F.L.S., F.Z.S., M.B.0.U. PART XIX.

Descrizione.— Maschio e femmina adulti.— Molto simile all'auritum, ma presenta parti superiori grigio cenere scuro, quasi nero sul collo anziché grigio-blu scuro, mentre il groppone è molto più chiaro. rispetto a quella specie. Quest'ultima mostra anche il colore delle parti superiori praticamente uniforme dalla nuca alle copritrici caudali superiori. Nel Crossoptilon a. harmani il bianco sulle guance e auricolari si estende in un'ampia fascia bianca attraverso la nuca, mentre auritum mostra solo una debole indicazione di bianco sulla nuca; nella prima razza anche il bianco del mento si espande in una linea sottile lungo la gola e collo.
Lati di quest'ultimo e parte superiore del petto grigio cenere molto scuro e lucido, che gradualmente vira in grigio cenere più chiaro sui fianchi e sulla parte inferiore del petto e bianco al centro dell'addome. Coda blu-nero metallico, con riflessi verdi e blu, mentre le penne centrali virano in grigio violaceo alla base. Non c'è bianco sulle timoniere esterne come nel Fagiano orecchiuto di Pallas.
Colori delle parti molli: maschio, lunghezza 89,53 cm.; iride marrone-arancio; becco rosso-corneo chiaro; zampe scarlatte; Gora 3048 metri (Cap. F. M. Bailey). La femmina, fatta eccezione per la lunghezza indicata di 86,68 cm, è descritta dal Capitano Bailey come avente esattamente le stesse parti molli colorate. In un altro maschio, apparentemente giovane, il Capitano Bailey descrive il colore dell'iride come marrone.
Misure: lunghezza totale da 81,28 a 93,98 cm. Becco anteriore lungo circa 41,9 mm., e dalla bocca 45,7 mm.; tarso da 87,6 a 92,7 mm.; ali da 292,1 a 321,3 mm.; coda da 457,2 a 558,8 mm. Gli speroni in un maschio di buona qualità misurano oltre 19,0 mm., e sembrano essere dello stesso colore delle zampe, con punte più scure.
Pulcino esaminato, probabilmente di circa due settimane di vita.— Corona nera vellutata e non marrone come nel Fagiano orecchiuto di Pallas; piumaggio superiore nero opaco che vira in grigio cenere scuro sul groppone e copritrici caudali superiori; spalle e ali vermicolate con barre rossastre con le copritrici alari che presentano ampie striature fulve rossastre. La distribuzione del bianco sulla testa è la stessa dell'adulto e i lunghi ciuffi auricolari sono parzialmente sviluppati, ma il bianco non si estende lungo la gola; è presente una macchia nera ben definita dietro e sotto le copritrici auricolari.
La parte superiore del petto e fianchi presenta piume nere con il centro e bordi terminali fulvi; parte inferiore del petto e addome bianco sporco; copritrici caudali e area anale grigio cenere opaco con apici bianchi; timoniere blu-nere con iridescenze blu, che assume una sfumatura verde in determinate condizioni di luce.
Giovane.— Iride marrone; becco color corno, più chiaro nella parte inferiore; zampe marrone rossastro (F. M. Bailey). L'ala di questo soggetto è lunga circa 177,8 mm.
Distribuzione: colline di Abor e Mishmi e Tibet sud-orientale. Apparentemente comune su tutte le catene montuose più elevate dell'estremo bacino idrografico nord-orientale del fiume Brahmaputra.
Nidificazione: sconosciuta. Il piccolo descritto sopra è stato catturato da Bailey il 16 luglio, e un altro piccolo "ben cresciuto" il 24 agosto, mentre nidiate di piccoli appena nati sono state osservate il 18 luglio.
Questo sembrerebbe indicare che deponevano le uova negli ultimi giorni di maggio, giugno e forse luglio.
Evidentemente si riproducono ad altitudini molto elevate, dai 3.000 metri in su, e la presenza effettiva di neve non può essere un deterrente per la loro nidificazione e schiusa.
Abitudini generali: per molto tempo l'unico esemplare conosciuto di questa forma è stata una pelle molto imperfetta ottenuta dal signor Elwes da un certo tenente Harman, che a sua volta l'aveva ricevuta da uno dei suoi periti indigeni. Disse di averla procurata a 250 chilometri a est di Lhasa, a un'altitudine di circa 1800 metri, dove si riuniva in stormi durante l'inverno. Questo avvenne nel 1881 e non si seppe più nulla della specie, delle sue abitudini e del suo habitat fino al 1913, quando il Capitano F. M. Bailey procurò una preziosa, seppur piccola, collezione di pelli di uccelli, tra le quali una splendida serie di fagiani di Elwes, contenente stupendi esemplari di maschi, femmine e giovani.
Nel Bombay Natural History Journal, il Capitano Bailey fornisce un resoconto molto interessante delle abitudini di questo splendido uccello:
"Questa specie si trova a Po Me, dove ho trovato piume e terreno razzolato, anche se non sono mai stato abbastanza fortunato da incontrare gli uccelli stessi. È comune nella bassa valle dello Tsangpo in Tibet. Il punto più a ovest in cui lo abbiamo visto era il versante orientale, Putrang La, dove ce n'erano molti nella macchia di rododendri a circa 4.724 metri di altitudine. La quota più bassa a cui abbiamo trovato questo fagiano nella valle dello Tsangpo era a circa 2.835 metri a Gyala, ma ne ho visto tracce a Po Me a circa 2.590 metri. Nella valle dello Tsangpo, il punto più alto a monte in cui li abbiamo trovati è stato nei pressi di Nang Dzong. Erano abbondanti nella valle del Char, un ramo del Subansiri, ma nessuno è stato trovato a ovest del Pu La, cioè lo spartiacque tra lo Tsang Po e il Subansiri
in questa regione. Ce n'erano molti sul Takarla e Le-La. Sono stati uditi emettere richiami vicino a Natrampa, nella bassa valle di Chayul, ma non sembrano estendersi a ovest di questi luoghi. Si diceva che fossero comuni a Tsari in inverno, ma non ne abbiamo visti. Si muovono in stormi di circa 5 o 10 esemplari frequentando le colline dominate da foreste e, alle quote più elevate, la giungla di rododendri nani, dove si nutrono nelle radure erbose tra i cespugli. Sono molto rumorosi al mattino presto e meno la sera. Il loro richiamo è simile a quello del C. tibetanum.
Quando sono allarmati, di solito si rifugiano sui rami degli alberi; il volo è pesante e solitamente in discesa. I battitori, vedendo questi uccelli, emettono un rumore simile all'abbaiare dei cani, gli uccelli spaventati cercano riparo su qualche albero e nel mentre vengono facilmente colpiti. Dicono che se non emettono questo suono, i fagiani orecchiuti volano a distanze considerevoli. Covate di pulcini appena nati sono state viste a Gyala, a 3048 metri di altitudine, il 18 luglio, mentre più o meno nello stesso periodo sono stati catturati dai tibetani nella bassa valle di Tsangpo un esemplare di "pulcino più grande", ed un giovane ben cresciuto sul Putrang La il 24 agosto. Esemplari adulti "in carne e ossa" misuravano: maschi 79,37, 85,72, 89,53 cm., e femmine 86,36 e 86,86 cm.
In una lettera a me indirizzata, il Capitano Bailey aggiunge alcune note interessanti. Afferma che, sebbene gli uccelli emettessero richiami principalmente al mattino, e in misura minore durante il ritorno al posatoio la sera, potevano essere uditi anche a intervalli irregolari durante il giorno, tranne forse le due o tre ore più calde del pomeriggio. Si tenevano molto vicini alla foresta avventurandosi negli spazi aperti solo al mattino e sera, ma, anche in quel caso non sembravano allontanarsi molto, ed al minimo allarme si rifugiavano tra i cespugli.
Sembra che abbia scoperto che sia il maschio che la femmina si prendevano cura dei piccoli, il che dimostrerebbe che i fagiani orecchiuti praticano la monogamia.
CROSSOPTILON MANTCHURICUM Swinhoe
Crossoptilon tibetanum.— Lamprey, (nec Hodgs.) Proc. Zool. Soc., 1862, p. 221. Crossoptilon auritum (sive = o oppure, ecc.) mantchuricum.— Swinh., Proc. Zool. Soc., 1862, p. 286; 1863, p. 306. Crossoptilon auritum.— Sclat., List of Phasianide, p. 6, pl. 9 (1863) ; Milne-Edwards, Nouv. Arch. Mus. Bull. L., p, 12, pl. 1, figs. 1 and 2 (1865); Sclat., Proc. Zool. Soc., 1866, p. 418; Saurin, Proc. Zool. Soc., 1866, p. 437; David, Nouv. Arch. Mus. Bull. Tll., p. 37 (1867); Bartlett, Proe. Zool. Sec., 1868, p. lis Gould, Birds of Asia, VII., pl. 22 (1870).
Crossoptilon mantchuricum.— Newton, Ibis (1865) p. 861. David, Nouv. Arch. Mus. Bull. VIL, p. 11 (1871); Swinhoe, Pro. Zool. Soc., 1871, p. 399; Sclater, Proc. Zool. Soc. (1871) p. 495; Elliot, Mon. Phasianidee I., pl. 16 (1872); David and Oustalet, Ois. Chine, p. 405, pl. 106 (1877); Sclater, Proc. Zool. Soc., 1879, p. 118, pl. VIII, fig. 5 (egg); Ogilvie-Grant, Cat. Birds B. M. XXIL., p. 294 (1893); Dresser, Manual Pal. Birds, p. 672 (1903) ; Ingram, Ibis (1909), p. 462. Crossoptilum manchuricum.— Ogilvie-Grant, Hand-L. Game Birds, I., p. 254 (1895). Sharpe, Hand-L., I., p. 35 (1898) ; Oates, Cat, Higgs, B. Mi) 1p. 038: (1901).
Nomi vernacolari.—Hoké or Hoky (Chinese). Ho-chi (vide Saurin.)
Descrizione.— Maschio e femmina adulti.— Corona marrone-nera vellutata; parte superiore del dorso, ali intere, collo e petto marroni, scuri e quasi neri vicino alla testa, che gradualmente sbiadiscono posteriormente sulle ali e dorso. Allo stesso modo, nella parte inferiore il petto cambia da un marrone intenso e profondo vicino al collo a marrone cenere chiaro sull'addome e copritrici sottocaudali. Parte inferiore del dorso, groppone e copritrici sopracaudali in bianco cenere sporco. Mento, gola e copritrici auricolari allungate bianche, generalmente macchiate sulla gola di giallognolo. Timoniere centrali bianche con steli ed apici neri, questi ultimi con iridescenze purpuree. In ogni coppia successiva di remiganti il bianco diminuisce di estensione tanto che che sotto alla coda viene gradualmente sostituito da marrone porpora o nero, con solo i lunghi pennacchi disintegrati delle remiganti centrali che brillano di bianco.
È interessante notare che una femmina in possesso di Lord Rothschild al Tring Museum presenta una piccola quantità di bianco sulla seconda e terza primaria vicino alle basi dei vessilli esterni.
Colori delle parti molli: lo spazio nudo che circonda gli occhi è rosso scarlatto brillante, il becco è rossastro e corneo, con la punta scura e più rosso alla base vicino alle narici; zampe rosse; gli artigli sono rossastri e corneo.
I maschi hanno uno sperone corto e smussato lungo circa 10 cm, rossastro alla base e marrone corneo all'estremità.
Misure: becco anteriore circa 43,2 mm, apertura della bocca circa 50,8 mm, o poco meno, copritrici auricolari allungate da 44,4 mm a 63,5 mm; tarso 88,9 mm a 96,5 mm; coda, penne centrali 660 mm; generalmente da circa 559 mm a 609,6 mm; penne più esterne da circa 190,5 mm a 216 mm; ali tra 298,4 mm e 230 mm.
Pulcino: testa fulva, centro della corona castano con due strisce coronali marrone scuro e nere poco definite; una striscia marrone scuro dalla parte posteriore dell'occhio attraversa le copritrici auricolari fino alla nuca. Sopra castano indistintamente barrato di nero e con due ampie bande longitudinali bianco sporco; ali marrone pallido opaco più o meno screziate e barrate di bianco e fulvo. Parte inferiore grigio pallido opaco; fianchi e cosce macchiati di marrone pallido e fulvo opaco.
Distribuzione: Monti della Manciuria nella Cina nord-orientale.
È importante notare che, sebbene la pelle originale ricevuta da Swinhoe provenisse dalla Manciuria 1*), da allora non ne sono state ottenute altre, e i collezionisti di Owston non sono riusciti a procurarsene alcuna nel 1908.
Nidificazione. — Misselbrook registra la riproduzione di questi fagiani come segue: "Vengono deposte da 12 a 16 uova per covata; l'incubazione è di circa 28 o 30 giorni. Non ho visto i maschi prendere parte all'incubazione. La particolarità dei piccoli è l'estrema docilità; non mostrando la benché minima timidezza, più o meno come i polli domestici. Le uva sono di un colore pietra pallido uniforme e misura circa 5,84 x 4,31 cm.”
Ci sono tre uova nella collezione del British Museum, tutte deposte in cattività. La consistenza di queste è molto fine e compatta, con una lucentezza decisa ma tenue e una superficie liscia come quella delle uova d'anatra, ma in una delle tre sono presenti le stesse piccole punteggiature già notate nelle uova di altre specie comprese in questo genere. Il colore varia da grigio oliva molto pallido a camoscio sporco chiaro.
Due uova della mia collezione, donatemi dal signor H. Munt e deposte nel Giardino Zoologico di Londra, corrispondono bene a quanto sopra, ma sono ancora più simili a quelle delle anatre per consistenza, colore e forma. Le cinque uova variano in lunghezza tra 52,1 mm., e 57,1 mm., e in larghezza tra 38,4 mm., e 39,9 mm.; come le uova di tutti i fagiani orecchiuti, sono molto piccole in proporzione alle dimensioni di chi le ha deposte.
Abitudini. — Pére David (Oiseaux de la Chine) scrive di questo uccello: "Il Crossoptilon bruno, noto a Pechino come Hoky, è residente in alcune zone boscose dei monti Péchély, ma da alcuni anni è diventato molto raro e, a causa della guerra di sterminio che gli viene condotta e della distruzione dei suoi habitat boschivi, non passerà molto tempo prima che scompaia del tutto. È una specie molto docile e socievole, sempre visibile in stormi. Si nutre con ogni tipo di semi, germogli, foglie, radici e insetti. Sembra essere ben adatto all'addomesticamento, soprattutto perché è così facile da nutrire, ma sarebbe necessario un parco ombreggiato nelle vicinanze di un ruscello d'acqua limpida; vale a dire condizioni simili a quelle che lo circondano allo stato selvatico."
Ancora lo stesso naturalista osserva (Nouv. Arch. de Mus. Bull.); "Una cosa che mi colpì una volta, quando a luglio ho ucciso due maschi e una femmina, tutti adulti, trovai quattro adulti e quindici giovani tutti insieme in uno stormo. Erano due famiglie ?. Questi uccelli si appollaiano volontariamente e tengono la coda disposta a forma di tetto (cioè compressa) e sollevata in alto come in un normale pollo."
Immagini, note e altro ancora, di specie non più riconosciute, apparenti ibridi e alcuni "strani" fenotipi osservabili nelle aree, più o meno, intermedie tra Crossoptilon c. crossoptilon, harmani, likiangense, mantchuricum e, probabilmente, auritum. In riguardo ai Crossoptilon c. harmani e dolani, altre immagini, definibili emblematiche, sono già allegate alla parte dedicata a questa ultima forma.
Crossoptilon leucurum (Fagiano orecchiuto dalla coda bianca) non riconosciuto ed incluso in Crossoptilon c. drouyini
Crossoptilon leucurum, Seebohm, Bull. Brit. Orn. Club, No. iv. p- xvii. (1892) ; Ogilvie-Grant, Cat. B. Brit. Mus. xxii. p. 294 (1893).
Maschio adulto —— Si distingue da Crossoptilon tibetanum per la maggior parte delle remiganti della coda bianche, tutte di un bianco puro, con estremità nere lucide di blu-violaceo.
Femmina adulta —— Ha il bianco sulle remiganti della coda meno esteso, e le coppie mediane ed esterne hanno le membrane interne grigie, mentre tutte sono con la punta e i margini grigio scuro.
Area di distribuzione: Tibet orientale; si incontra tra il Passo Sok e Chiamdo, anche sull'altopiano tra il Passo Sok e Lhassa.
Gli esempi tipici di questa specie apparentemente perfettamente distinta furono ottenuti dal Capitano Bower e dal Dott. Thorold tra il Passo Sok e Chiamdo, e esemplari simili furono raccolti dal Prince Henry of Orleans e da M. Bonvalot alcuni anni prima sull'altopiano tra il Passo Sok e e Lhassa. Questi ultimi uccelli sono ora conservati al Museo di Parigi e il Dr. Oustalet li considera semplicemente varietà di Crossptilon tibetanum, o ibridi tra questa specie e il Crossoptilon auritum, Pallas, di colore grigio ardesia, le cui penne esterne della coda sono per la maggior parte bianche. Siamo pienamente d'accordo con il signor Seebohm nel ritenere che questa conclusione sia errata, poiché il Crossoptilon auritum, possiamo ulteriormente osservare, ha la coda composta da ventiquattro o venti penne. Sembra che l'areale del Crossoptilon leucurum si sovrapponga a quello del Crossoptilon tibetanum nel Tibet orientale, ed è possibile che in questa località le due forme si incrocino, quindi è possibile che il tipo di Crossoptilon tibetanum di Hodgson, che presenta alcune macchie bianche sulle sei paia esterne di penne della coda possa, possa essere un uccello incrociato di questa descrizione, ma la località esatta in cui è stato ottenuto è piuttosto incerta.
%2C%20Lhorong%20prefettura%20di%20Qamdo%2016-05-24%20-%20.jpg)
Crossoptilon leucurum - www.douyin.com Autore: Neve a Sabei ( località indicata dal fotografo: Contea di Lhorong prefettura di Qamdo, Tibet 16-05-2024 Cina )
Nota: perlomeno da quello che "raccontano" le immagini e video, anche molto recenti, esemplari che mostrano questa colorazione oltre ad essere molto uniformi nel fenotipo, sicuramente molto più di conspecifici ritenuti specificatamente distinti, sembrerebbero rappresentati da popolazioni piuttosto numerose.
Foto da: www.douyin.com Autore: Neve a Sabei ( località indicata dal fotografo: Contea di Lhorong prefettura di Qamdo, Tibet 16-05-2024 Cina )
1 - Crossoptilon leucurum 2 - Crossoptilon leucurum
%2C%20Lhorong%20prefettura%20di%20Qa.jpg)
%2C%20%20citt%C3%A0%20a%20livello%20di%20pref.jpg)
Crossoptilon crossptilon specie con piumaggio arruffato
Foto da: 1 www.ebird.org Autore: Zhen Ting Li - 2-3 www.igoterra.com Autore: 2 Pauk Matson ( località indicata dai fotografi: 1 Yading National Nature Reserve [general area], Sichuan 27-05-2023 Cina - 2 (Baizha Forest), prefettura di Yushu, Qinghai 22-06-2025 Cina - 3 villaggio di Yiala o Yela, prefettura di Yushu, Qinghai, China 25-06-2025 Cina )
1 - Crossoptilon c. crossoptilon 2 - Crossoptilon c. drouyni 3 - Crossoptilon c. dolani
![Nominale con piuma riccia - (Yading NNR [general area]), Sichuan 27-05-23 - Zhen Ting Li e](https://static.wixstatic.com/media/3ebe19_320227b69ac242b481cd549c09ec3b75~mv2.jpg/v1/crop/x_299,y_69,w_739,h_699/fill/w_311,h_294,al_c,q_80,usm_0.66_1.00_0.01,enc_avif,quality_auto/Nominale%20con%20piuma%20riccia%20-%20(Yading%20NNR%20%5Bgeneral%20area%5D)%2C%20Sichuan%2027-05-23%20-%20Zhen%20Ting%20Li%20e.jpg)


Crossoptilon crossptilon specie con piumaggio arruffato
Nota: per la foto 1 l'identificazione è presunta in base al fenotipo mostrato ed alla, seppur minima, indicazione di località. Nella 2 un soggetto con coda pressoché solo bianca, presumibilmente uno dei tanti ibridi/intermedi osservabili in queste aree.
Foto da: 1 Autore: Lao Photograpphy - 2 Zongzhuang Liu ( località indicata dai fotografi: 1 solo Yunnan 03-05-2025 Cina - 2 (Baizha Forest), prefettura di Yushu, Qinghai 25-06-2025 Cina )
1 - Crossoptilon c. lichiangense (probabile) 2 - Crossoptilon c. drouyni/leucurum
%20(1)_e.jpg)
%2C%20Qinghai%2C%20China%20%2025-06-25%20-%20Zongzhuang%20Liu.jpg)
Crossoptilon mantchuricum con fenotipo, apparentemente, anomalo: mutazione o ibrido ? (simile, per alcuni aspetti, agli ibridi menzionati da Ghigi nel suo lavoro, di ricerca ibridologica riguardante i Crossoptilon, del 1934 esposto a seguito)
Nota: nella serie sottoesposta (6 foto) con esemplari di Crossoptilon mantchuricum, groppa e copritrici della coda sembrano vistosamente grigiastri anziché essere bianco sporco come di norma. Certo, l'ambiente circostante appare come brunastro/grigiastro, quindi potrebbe favorire l'illusione visiva della colorazione grigia. Tuttavia, per vari motivi, come ad esempio posture assunte nel momento degli scatti fotografici, le diverse condizioni di luce così come il palesarsi del bianco sulle timoniere più esterne oppure il grigiastro con rachidi bianchi della coda vista da sotto (foto 2), parrebbero escludere che possa trattarsi di un effetto ottico. Per cui, se così fosse...., mutazione o ibrido ?
Foto da: 1-2-3 www.birdnet.cn Autore: Gushan ( località indicata dal fotografo: putroppo come spesso accade con i fotografi cinesi, non è stato menzionato alcun luogo. Tuttavia, considerato il tipo di specie in oggetto, ciò è di relativa importanza )
1 - Crossoptilon mantchuricum 2 - Crossoptilon mantchuricum 3 - Crossoptilon mantchuricum



Crossoptilon mantchuricum con fenotipo, apparentemente, anomalo: mutazione o ibrido ?
4 - Crossoptilon mantchuricum 5 - Crossoptilon mantchuricum 6 - Crossoptilon mantchuricum

.jpg)

Da: L'OISEAU - ET LA REVUE FRANÇAISE D'ORNITHOLOGIE - Volume IV — Nouvelle Série — Année 1934
Sintesi degli studi compiuti da Ghigi e Delacour
RICERCA IBRIDOLOGICA SUI CROSSOPTILON
di Alessandro Ghigi
Durante l'estate del 1929, il signor Jean Delacour importò dalla Cina una notevole quantità di Crossoptilon auritum vivi, quasi tutti maschi. Me ne diede due, uno dei quali, accoppiato con una femmina di C. mantchuricum, produsse venti pulcini nel 1930; riuscii ad allevarne quindici.
"..Nel 1932, ottenni alcuni esemplari F2 e diversi esemplari dell'incrocio (auritum x mantchuricum) x mantchuricum, mentre il signor Delacour allevò 12 esemplari dell'incrocio auritum x (auritum x mantchuricum).
Tutto il materiale a disposizione di Delacour e mio consente ora uno studio ibridologico di queste forme di Crossoptilon. Da un punto di vista morfologico, non vi sono differenze apprezzabili tra queste due forme selvatiche, fatta eccezione per le timoniere mediane, che sono completamente decomposte in C. auritum, mentre in C. mantchuricum le barbe sono normali e compatte all'apice. Inoltre, la coda di C. mantchuricum è notevolmente più lunga di quella di C. auritum.
Il colore generale del corpo di C. auritum è blu ardesia, da cui il nome Blue Hoki; C. mantchuricum, invece, è marrone, con dorso e groppone bianco sporco.
Questa parte del corpo presenta quindi due gruppi di caratteristiche antagoniste:
1. Omogeneità (auritum) ed eterogeneità (mantchuricum) del colore, rispetto alle altre parti del corpo;
2. Sfumatura blu (auritum) e tinta bianca (mantchuricum).
Sulle timoniere laterali della coda si possono osservare tre caratteristiche opposte:
1. In auritum, la porzione basale bianca è nettamente distinta dalla parte terminale nera, mentre in mantchuricum, la transizione dal bianco al marrone scuro è attenuata;
2. In aurilun, la parte basale è bianco puro; mentre in mantchuricum, il bianco è sporco;
3. In auritum, la porzione apicale è nero acciaio con riflessi verdi, mentre in mantchuricum, la porzione corrispondente è nero-brunastra, con riflessi violacei poco brillanti.
In conclusione, le seguenti differenze influenzano le seguenti caratteristiche:
1. Struttura delle barbe all'apice delle timoniere mediane 2. Colore delle timoniere mediane. 3. Lunghezza della coda. 4° Colore delle timoniere laterali delle coppie 3-6. 5° Colore della base delle 5 paia di timoniere esterne. 6° Colore della metà apicale delle 5 paia di timoniere esterne. 7° Separazione tra la metà bianca e quella scura delle stesse timoniere. 8° Uniformità o eterogeneità di colore tra il groppone e il resto del corpo. 9° Colore del groppone. 10° Colore generale del corpo:
1. Nelle penne mediane della coda, come in mantchuricum, è presente una porzione apicale compatta; è lunga 60-70 mm e larga 20-25 mm. Come in mantchuricum, la sua lunghezza è di 95-120 mm e la sua larghezza di 30-35 mm; mentre in auritum le barbe sono completamente decomposte; l'ibrido è, sotto questo aspetto, intermedio.
2. Le penne mediane della coda in C. auritum sono blu scuro, mentre in mantchuricum l'intera parte decomposta è bianco sporco; nell'ibrido F1, l'intera penna è grigia, meno scura che in auritum, ma più simile.
3. La lunghezza della coda nell'ibrido è di 360-390 mm; in auritum è di 350 mm e in manchuricum di 400 mm; pertanto, gli ibridi sono intermedi e variabili.
4° Il colore delle timoniere laterali del sesto paio è, in auritum, blu-nero, con riflessi verdi brillanti, ad eccezione di una linea bianca irregolare sul sesto paio; in manchuricum, tre quarti delle penne sono biancastre alla base, mentre la punta è bruno-violacea; nell'ibrido, la base è grigia e la tonalità vira gradualmente al blu-nero con riflessi acciaio; vi è quindi una dominanza, seppur incompleta, del carattere di auritum.
5° Il colore della parte basale delle 5 paia di timoniere esterne è bianco sporco in C. manchuricum, bianco puro in C. auritum. Quest'ultimo carattere è dominante nell'ibrido.
6° Il colore della parte apicale delle stesse timoniere è nero, sfumando al blu-verde in C. auritum; è grigio-marrone sfumando al violaceo in mantehuricum; nell'ibrido, il carattere auritum è dominante.
7° Nell'auritum i due colori sono nettamente separati, per cui alla base della coda è presente una macchia bianca; nel mantchuricum la transizione dal bianco al grigio-marrone è graduale, con tonalità intermedie; nell'ibrido è dominante il carattere auritum, per cui alla base della coda è presente una macchia bianca.
8° Il groppone e le copritrici della coda sono colorate come il resto del corpo in C. auritum; sono diverse e di un bianco sporco in mantchuricum; nell'ibrido, queste parti sono generalmente omogenee con il resto del corpo, tuttavia, in alcuni esemplari la tonalità è meno scura sulle copritrici della coda. L'omogeneità di auritum è quindi dominante, sebbene variabile e imperfetta.
9° Il groppone è blu in auritum, bianco sporco in mantchuricum; nell'ibrido è grigio, a volte più scuro o più intenso. Vi è quindi una dominanza, sebbene in qualche modo attenuata, della tonalità scura di auritum.
10° Il colore generale del corpo è blu in auritum, marrone in mantchuricum; nell'ibrido è un grigio intermedio, ma meno diverso da auritum che da mantchuricum.
F2 — C. auritum x C. mantchuricum
Da diverse coppie F, abbiamo ottenuto pochi esemplari nel 1932, sia perché alcune femmine non deponevano uova, sia perché la mortalità embrionale era molto elevata. Tuttavia, escludendo alcuni individui nati da altri riproduttori che non ho potuto esaminare, fornirò qui alcune informazioni su tre esemplari del 1932.
In generale, la separazione non è molto evidente e i giovani assomigliano molto ai genitori; rientrano almeno nell'intervallo delle loro varianti, soprattutto per quanto riguarda il colore delle copritrici della coda, che a volte sono più chiare di quelle del groppone. La macchiolina bianca alla base della coda è marcata come in F.I. e C. auritum. È da notare che nell'esemplare più scuro, le remiganti omerali, le scapolari e le copritrici alari minori sono ombreggiate di blu metallico, più evidenti che in auritum e su uno sfondo più scuro rispetto alle due specie genitori.
Pur riservandomi il diritto di fornire ulteriori informazioni qualora nell'anno in corso (1933) un numero maggiore di esemplari rivelasse qualche fatto nuovo, posso concludere che l’F2 assomiglia molto a F1 e che i fenomeni di disgiunzione tendono verso la forma auritum.
INCROCIO C. auritum x (C. auritum x C. mantchuricum)
Il signor Delacour ha allevato 12 esemplari, una coppia dei quali è ora in mio possesso.
Il colore del corpo è blu nella maggior parte degli esemplari, quindi non è possibile distinguerli dalla specie pura di auritum. Due esemplari presentano tonalità brunastre che li rendono intermedi tra l'ibrido F1 e l'auritum.
La porzione apicale della coda è decomposta nella maggior parte degli esemplari, ma ce ne sono alcuni che la presentano normale, come nel mantchuricum. In questo caso, abbiamo un auritum con penne caudali mediane con una struttura simile a quella del mantchuricum.
Re-incrocio (C. auritum x C. mantchuricum) x C. mantehuricum
Ho allevato 12 individui dalla stessa coppia, composta da un maschio ibrido e una femmina di mantchuricum.
In generale, possono essere considerati intermedi tra F1 e mantchuricum, con una maggiore somiglianza con quest'ultima specie rispetto all'ibrido di prima generazione. Non sono uniformi nel colore del groppone e delle penne superiori della coda. Come in F1, abbiamo segnalato alcuni individui con penne superiori della coda più chiare delle penne del groppone; lo stesso fenomeno si osserva in questo incrocio; ci sono uccelli con la parte centrale del groppone scura come il dorso.
Il colore generale del corpo è quello di mantchuricum, il colore della coda invece, ad eccezione di una tonalità più chiara propria di questa specie, sulle timoniere mediane, assomiglia maggiormente a F1 e di conseguenza a auritum, soprattutto per la brillantezza dello specchio bianco alla base delle timoniere esterne.
Un nuovo carattere appare sulle penne superiori della coda: queste piume, nei genitori di entrambe le specie, sono uniformemente bianco sporco nel mantchuricum e blu nell'auritum; inoltre, nell'ibrido F1 sono uniformemente grigio scuro o grigio chiaro, a seconda dei soggetti. In molti di questi incroci, si alternano strisce chiare e scure, che conferiscono all'uccello un carattere molto particolare.
Confrontando questi due incroci, si scopre che i tratti di colore di auritum sono dominanti nel primo, mentre quelli di mantchuricum non sono completamente dominanti nel secondo. In quest'ultimo, osserviamo una disgiunzione e la comparsa di un nuovo tratto sul groppone: la striscia.
Da una prospettiva genetica generale, osserviamo che la prima generazione F1 non è uniforme; che negli incroci si verifica una disgiunzione di alcuni tratti e che il colore della specie auritum si comporta come un tratto dominante, mentre quello della specie mantchuricum non lo è altrettanto.
Ora, cercherò di discutere l'importanza di questa ricerca sperimentale per l'intera sistematica del genere Crossoptilon. Tutti gli autori concordano nel considerare auritum e mantchuricum due buone specie, sebbene appartengano alla sezione con 22-24 timoniere, le quattro centrali delle quali presentano barbe decomposte, con ciuffi auricolari che si estendono oltre le penne della testa. Queste due specie sono in contrasto con Crossoptilon crossoptilon (=tibetanum), che presenta 20 timoniere, tutte con barba normale, e ciuffi auricolari corti, che non si estendono oltre l'occipite.
Da un punto di vista fisiologico, appartengono alla stessa specie naturale e i risultati dell'esperimento hanno dimostrato che la loro esistenza è legata all'isolamento geografico. Se le due forme non abitassero due territori molto distanti e se, al contrario, avessero contatti lungo i loro areali di distribuzione, assisteremmo a tutta una serie di intermediari, con prevalenza però della forma auritum, la cui dominanza appare soprattutto nei reincroci, mentre la forma mantchuricum non riappare nella sua interezza.
Il signor W. Beebe, nella sua monografia sui fagiani, ha espresso l'opinione che C. harmani Elwes sia un ibrido tra C. crossoptilon e C. auritum. Il dottor Hartert e Lord Rothschild hanno osservato che in nessuna regione convivono due diverse forme di Crossoptilon e che, in ogni caso, le tre che ho appena menzionato abitano aree geografiche molto distanti tra loro. Lord Rothschild ritiene che C. crossoptilon (=tibetanum) sia divisibile in tre sottospecie: C. crossoptilon crossoptilon, C. c. leucurum e C. ce. harmani, le prime due più o meno bianche, la terza blu ardesia. Quando ottenni i miei primi ibridi auritum x mantchuricum, volevo esaminare gli esemplari di Crossoptilon presenti al Museo di Storia Naturale di Parigi, assistito dal mio competentissimo amico, il signor Delacour, il quale, con la sua consueta cortesia, chiese gentilmente alcune informazioni a Lord Rothschild dopo la nostra visita al Museo. Possedeva una ricchissima collezione di questi uccelli nel suo museo di Tring.
Ho ricevuto, tramite il signor Delacour, tre lettere da Lord Rothschild. Il 16 novembre 1930, ci avvertì che auritum e mantchuricum non hanno nulla a che fare con crossoptilon-tibetanum, perché le prime due specie presentano un maggior numero di penne caudali, le cui mediane sono completamente decomposte; pertanto, l'opinione di Beebe secondo cui C. harmani sia un ibrido tra auritum e crossoptilon è inammissibile. Scrive inoltre di aver ricevuto due esemplari da Parigi, uno dei quali raccolto dal principe H. d'Orléans vicino a Djala, nel Tibet sud-orientale; Possiede ancora una serie di dieci esemplari raccolti da Forrest a Lichiang, nelle montagne dello Yunnan nord-occidentale, una serie che può essere descritta.
L'esemplare di Djala, proveniente dall'8° est del Tibet, è bianco puro, fatta eccezione per la parte superiore della testa, le primarie marrone scuro e il terzo basale della coda anch'esso bianco puro; egli ritiene che questo esemplare rappresenti C. leucurum. Ciò che confonde la questione è che Forrest ha raccolto giovani e da 12 a 15 adulti di entrambe le specie, che non sono bianchi come l'esemplare del Principe d'Orléans; sono leggermente più grigio-crema, e il terzo basale della coda è grigio scuro, quasi fuligginoso; le primarie sono grigio scuro, quasi nerastre, meno scure all'esterno, con rachide nero. In breve, potrebbero appartenere a drouyni; in questo caso, avremmo un'ulteriore razza locale, che abita un'area definita, dove tutti gli esemplari sono costanti, ma ai confini della razza più vicina incontriamo ogni sorta di individui intermedi. In una seconda lettera del 17 novembre 1930, Lord Rothschild ci informa di aver esaminato la serie di Crossoptilon crossoptilon del British Museum, tra cui i tipi di C. e. leucurum e C. c. crossoptilon. Il primo è bianco puro, con rachide bianco delle primarie e solo 6,5-7,5 cm di nero sulla punta della coda; tuttavia, esistono altri esemplari della stessa località e collezione, con molto meno bianco sulla coda e rachide nero delle primarie. Il tipo di C. c. crossoptilon è molto più grigio-argenteo sulle copritrici alari ed è meno bianco puro; inoltre, tutte le remiganti, con i loro rachide, sono uniformemente bruno-nero fuligginoso e la coda completamente scura. Gli esemplari di Forrest del British Museum (5 o 6) mostrano ogni sorta di variazione delle primarie, dal grigio al marrone scuro uniforme. Lord Rothschild conclude che esistono tre razze locali di C. crossoptilon:
C. c. crossoptilon (Hodgs.) del Sichuan occidentale. C. c. leucurum ( Seeb.) del sud-est Tibet. C. c. harmani ( Elwes ) del Tibet sud occidentale
Tuttavia, tutte le gradazioni di colore si verificano ai confini delle aree di dispersione delle tre razze. Il tipo di C. c. leucurum, della collezione Seebohm, reca l'iscrizione: S.-E. Tibet.
Lord Rothschild mi ha anche gentilmente inviato una fotografia di un ibrido tra C. mantchuricum e C. crossoptilon, ottenuto presso il Giardino Zoologico di Anversa. In questo esemplare la coda è normale, ovvero il carattere delle barbe decomposte è recessivo. Il groppone è biancastro con la base di ogni penna grigia, un tratto comune a entrambe le specie parentali. Le altre parti del corpo sono di colore intermedio: petto e ventre sono grigio chiaro, risultato della combinazione del bianco di crossoptilon con il marrone di mantchuricum; dorso, scapolari e copritrici alari sono più scuri, perché non sono completamente bianchi in crossoptilon; le remiganti secondarie e primarie sono molto scure, come nella specie parentale. Le creste auricolari sono notevoli in quanto sporgono oltre le piccole penne della testa.
Ora, tenendo conto di tutta la serie dei nostri ibridi tra auritum e mantchuricum e di quella del Giardino di Anversa tra crossoptilon (tibetanum) e mantchuricum, esaminerò se harmani possa essere considerato, dal punto di vista genetico, un ibrido tra crossoptilon e auritum. Per quanto riguarda le timoniere decomposte, va notato che l'ibrido auritum x mantchuricum è intermedio, ovvero il carattere di auritum si fa sentire al punto da diminuire la parte normale della punta. Al contrario, in questo ibrido non sono presenti barbe decomposte, ma non conosciamo il numero di timoniere. Il colore di harmani potrebbe derivare da auritum, perché abbiamo visto che il colore scuro di questa specie è dominante sul bianco sporco di mantchuricum. Ma la caratteristica che, a mio avviso, risolve la questione sono i ciuffi auricolari, che sono notevolmente più lunghi di quelli di crossoptilon. Se harmani fosse un ibrido crossoptilon x auritum, dovrebbe avere lunghi ciuffi come l'ibrido crossoptilon x mantchuricum. Al contrario, possiede il carattere crossoptilon, recessivo nell'incrocio con una specie con lunghi ciuffi auricolari.
Gli esemplari del Museo di Parigi confermano i dati di Lord Rothschild: tutti i leucurum sono di colore bianco puro, ad eccezione delle punte delle timoniere, dei rachide delle primarie e della sommità della testa, che sono neri. Crossoptilon (tibetanum) presenta una peluria basale sulle timoniere posteriori, lati sempre più scure nelle parti coperte delle timoniere fino alle remiganti e timoniere caudali interamente blu con una sfumatura acciaio. C. harmani assomiglia all'auritum per colore, ma ha le parti inferiori bianche e una coda acciaio, macchiata di bianco lungo la parte interna della membrana esterna delle timoniere laterali.
Se volessimo rappresentare la gradazione delle tonalità in ordine lineare, dal bianco al blu e al marrone, dovremmo disporre la serie come segue:
leucurum (Seeb.) drouyni (Verr.) crossoptilon (Hodgs) harmani (Elwes.) auritum (Pall.) -- mantchuricum (Swinb.)
Le prime quattro forme sono ovviamente le più primitive: probabilmente la specie originaria è crossoptilon, che si è evoluta a ovest in leucurum e a est in harmani, mentre auritum e mantchuricum sono due forme più distinte e notevolmente divergenti. Infatti, nei giovani di queste ultime forme, quando sono nel loro primo piumaggio, sono presenti solo 20 penne della coda con barbe normali; la muta è centripeta come in tutti i fagiani; sono le penne laterali della coda a cambiare per prime. Quelle caudali mediane con barbe decomposte compaiono contemporaneamente alle laterali; rappresentano quindi un'acquisizione tardiva che non ha corrispondenza in altri fagiani. I ciuffi auricolari nei giovani non si estendono oltre la testa. Pertanto, in auritum e mantchuricum, sono presenti due fattori di sviluppo: penne mediane della coda decomposte e ciuffi auricolari lunghi, che non appartengono al gruppo crossoptilon-leucurum-harmanti.
Lo stato intermedio degli ibridi dimostra inoltre che auritum e mantchurieum si sono evoluti in uno stato di isolamento geografico, mentre crossoptilon è sempre stato in comunione geografica con i suoi stretti alleati, come dimostrato dai numerosi intermedi rinvenuti in natura, di cui drouyni è il più noto.
Speriamo di importare esemplari vivi di Crossoptilon crossoptilon (tibetanum); l'incrocio con auritum potrebbe risolvere i dubbi rimanenti sulle affinità naturali delle diverse forme di questi curiosi fagiani.
Appendice. — Nel 1933, ho allevato 6 esemplari di C. auritum x C. mantchuricum (F. auritum) e 20 esemplari dall'incrocio (C. auritum x C. mantchuricum) x C. mantchuricum).
Due esemplari del primo lotto sono decisamente blu: si è quindi verificata una netta disgiunzione in direzione dell'auritum: è presente anche un esemplare molto marrone, ma poiché il colore scuro si estende al dorso e al groppone, l'uccello, nel suo complesso, rimane un ibrido intermedio.
Il secondo lotto conferma quanto osservato in precedenza: gli uccelli sono marroni con un bianco-grigiastro più o meno esteso sul dorso e sul groppone; quest'ultimo, sempre meno scuro, ma mai bianco puro come nella specie C. mantchuricum. Inoltre, sono presenti quattro esemplari con striature distinte sul groppone.
M. Stefani (Avicultural Magazine 1933, p. 223) deplora, dal punto di vista pratico, l'ibridazione effettuata in Francia e in Italia, perché potrebbe rovinare la genetica del Crossoptilon auritum, proprio come il Fagiano di Lady Amherst è stato contaminato dal Fagiano dorato.
Dai nostri esperimenti, possiamo concludere che la specie maggiormente in pericolo per la perdita di purezza somatica a causa dell'ibridazione è il Crossoptilon mantchuricum, ma non Crossoptilon auritum.."